欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (2): 377-391.doi: 10.3724/SP.J.1006.2023.23021
殷芳冰1( ), 李雅楠1, 鲍建喜1, 马雅杰1, 秦文萱1, 王锐璞1, 龙艳1,2, 李金萍2, 董振营1,2,*(), 万向元1,2,*()
), 李雅楠1, 鲍建喜1, 马雅杰1, 秦文萱1, 王锐璞1, 龙艳1,2, 李金萍2, 董振营1,2,*(), 万向元1,2,*()
YIN Fang-Bing1(), LI Ya-Nan1, BAO Jian-Xi1, MA Ya-Jie1, QIN Wen-Xuan1, WANG Rui-Pu1, LONG Yan1,2, LI Jin-Ping2, DONG Zhen-Ying1,2,*(), WAN Xiang-Yuan1,2,*()
摘要:
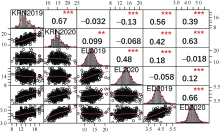
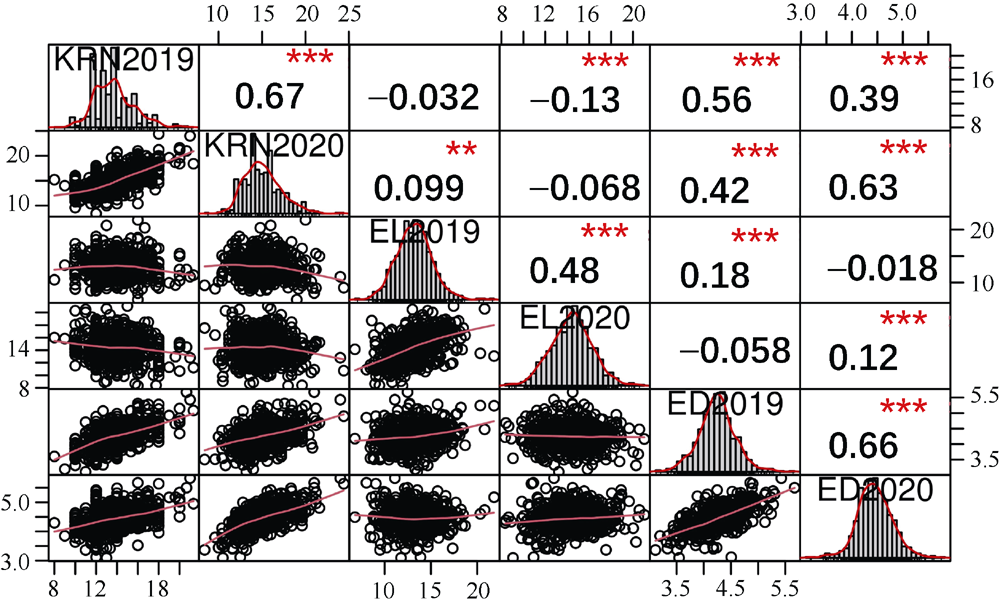
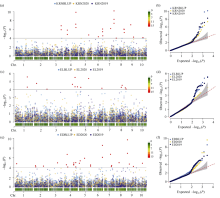
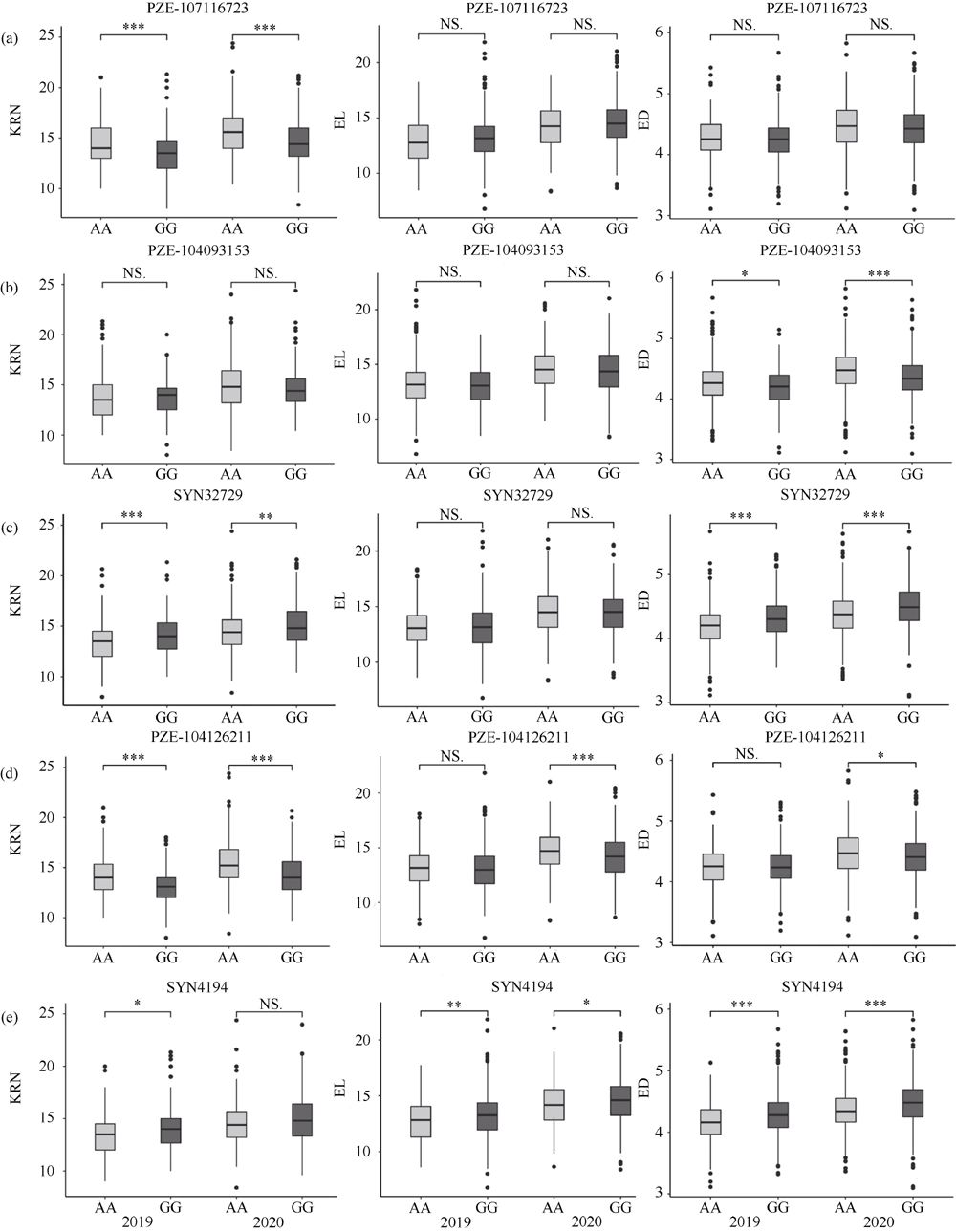
玉米雌穗产量相关性状直接影响玉米最终产量, 解析其遗传机制可为玉米高产提供有益指导。本研究以733份玉米自交系作为关联群体, 在2个环境下随机区组种植, 调查穗行数(KRN)、穗长(EL)和穗粗(ED) 3个产量相关性状, 利用MaizeSNP3072芯片对其进行基因分型, 采用FarmCPU模型进行全基因组关联分析, 分别鉴定出16、13和24个与3个性状显著关联的单核苷酸多态性位点(SNP), 对表型变异的解释率分别为0.01%~7.08%、0.01%~5.34%和0.07%~4.34%。其中, 分别有6、2和5个与3个性状存在显著关联的高可信度(high confidence, HC) SNP, 而且有2个HC-SNP同时与KRN和ED显著相关, 1个KRN HC-SNP和3个ED HC-SNP为本研究首次报道。在所鉴定HC-SNP上下游200 kb范围内筛选出33个重要候选基因, 其中9号染色体SNP标记PZE-109003046所在基因PIN1a为控制生长素极性运输从而调控雌穗性状的已知基因。另一些候选基因编码不同转录因子, 以及参与生长素、赤霉素和乙烯等激素介导的信号转导、DNA甲基化和蛋白磷酸化等翻译后修饰过程的蛋白, 可能从不同方面调控雌穗相关性状。本研究所挖掘的11个HC-SNP与33个候选基因可以为进一步克隆雌穗性状功能基因、揭示相关分子调控机制以及利用分子标记辅助选择育种提供有益指导。
| [1] | 宁慧云, 连晋, 赵玉坤, 连吉明, 高根来. 不同玉米品种雌穗性状及产量的灰关联评价研究. 农学学报, 2013, 3(6): 13-16. |
| Ning H Y, Lian J, Zhao Y K, Lian J M, Gao G L. Grey relational evaluation study in ear traits and yield of different maize varieties. J Agric, 2013, 3(6): 13-16. (in Chinese with English abstract) | |
| [2] | 王帮太, 吴建宇, 丁俊强, 席章营. 玉米产量及产量相关性状QTL的图谱整合. 作物学报, 2009, 35: 1836-1843. |
|
Wang B T, Wu J Y, Ding J Q, Xi Z Y. Map integration of QTLs for grain yield and its related traits in maize. Acta Agron Sin, 2009, 35: 1836-1843. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2009.01836 |
|
| [3] | 吴律, 代力强, 董青松, 施婷婷, 王丕武. 玉米行粒数的全基因组关联分析. 作物学报, 2017, 43: 1559-1564. |
|
Wu L, Dai L Q, Dong Q S, Shi T T, Wang P W. Genome-wide association analysis of kernel number per row in maize. Acta Agron Sin, 2017, 43: 1559-1564 (in Chinese with English abstract).
doi: 10.3724/SP.J.1006.2017.01569 |
|
| [4] |
张焕欣, 翁建峰, 张晓聪, 刘昌林, 雍洪军, 郝转芳, 李新海. 玉米穗行数全基因组关联分析. 作物学报, 2014, 40: 1-6.
doi: 10.3724/SP.J.1006.2014.00001 |
|
Zhang H X, Weng J F, Zhang X C, Liu C L, Yong H J, Hao Z F, Li X H. Genome-wide association analysis of kernel row number in maize. Acta Agron Sin, 2014, 40: 1-6. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2014.00001 |
|
| [5] |
Li F, Jia H T, Liu L, Zhang C X, Liu Z J, Zhang Z X. Quantitative trait loci mapping for kernel row number using chromosome segment substitution lines in maize. Genet Mol Res, 2014, 13: 1707-1716.
doi: 10.4238/2014.January.17.1 pmid: 24535896 |
| [6] |
Choi J K, Sa K J, Park D H, Lim S E, Ryu S H, Park J Y, Park K J, Rhee H I, Lee M, Lee J K. Construction of genetic linkage map and identification of QTLs related to agronomic traits in DH population of maize (Zea mays L.) using SSR markers. Genes Genomics, 2019, 41: 667-678.
doi: 10.1007/s13258-019-00813-x |
| [7] |
Zhou B, Zhou Z J, Ding J Q, Zhang X C, Mu C, Wu Y B, Gao J Y, Song Y X, Wang S W, Ma J L, Li X T, Wang R X, Xia Z L, Chen J F, Wu J Y. Combining three mapping strategies to reveal quantitative trait loci and candidate genes for maize ear length. Plant Genome, 2018, 11: 170107.
doi: 10.3835/plantgenome2017.11.0107 |
| [8] |
Zhang X X, Guan Z R, Li Z L, Liu P, Ma L L, Zhang Y C, Pan L, He S J, Zhang Y L, Li P, Ge F, Zou C Y, He Y C, Gao S B, Pan G T, Shen Y O. A combination of linkage mapping and GWAS brings new elements on the genetic basis of yield-related traits in maize across multiple environments. Theor Appl Genet, 2020, 133: 2881-2895.
doi: 10.1007/s00122-020-03639-4 pmid: 32594266 |
| [9] |
Xu Y, Xu C, Xu S. Prediction and association mapping of agronomic traits in maize using multiple omic data. Heredity, 2017, 119: 174-184.
doi: 10.1038/hdy.2017.27 pmid: 28590463 |
| [10] |
Li T, Qu J Z, Tian X K, Lao Y H, Wei N N, Wang Y H, Hao Y C, Zhang X H, Xue J Q, Xu S T. Identification of ear morphology genes in maize (Zea mays L.) using selective sweeps and association mapping. Front Genet, 2020, 11: 747.
doi: 10.3389/fgene.2020.00747 |
| [11] |
Tian H L, Wang F G, Zhao J R, Yi H M, Wang L, Wang R, Yang Y, Song W. Development of maizeSNP3072, a high-throughput compatible SNP array, for DNA fingerprinting identification of Chinese maize varieties. Mol Breed, 2015, 35: 136.
doi: 10.1007/s11032-015-0335-0 |
| [12] |
Zhu X M, Shao X Y, Pei Y H, Guo X M, Li J, Song X Y, Zhao M A. Genetic diversity and genome-wide association study of major ear quantitative traits using high-density SNPs in maize. Front Plant Sci, 2018, 9: 966.
doi: 10.3389/fpls.2018.00966 |
| [13] |
Zhang C S, Zhou Z Q, Yong H J, Zhang X C, Hao Z F, Zhang F J, Li M S, Zhang D G, Li X H, Wang Z H, Weng J F. Analysis of the genetic architecture of maize ear and grain morphological traits by combined linkage and association mapping. Theor Appl Genet, 2017, 130: 1011-1029.
doi: 10.1007/s00122-017-2867-7 pmid: 28215025 |
| [14] |
Xue Y D, Warburton M L, Sawkins M, Zhang X H, Setter T, Xu Y B, Grudloyma P, Gethi J, Ribaut J M, Li W C, Zhang X B, Zheng Y L, Yan J B. Genome-wide association analysis for nine agronomic traits in maize under well-watered and water-stressed conditions. Theor Appl Genet, 2013, 126: 2587-2596.
doi: 10.1007/s00122-013-2158-x pmid: 23884600 |
| [15] |
Pandis N. Linear regression. Am J Orthod Dentofacial Orthop, 2016, 149: 431-434.
doi: 10.1016/j.ajodo.2015.11.019 |
| [16] |
Carraro N, Forestan C, Canova S, Traas J, Varotto S. ZmPIN1a and ZmPIN1b encode two novel putative candidates for polar auxin transport and plant architecture determination of maize. Plant Physiol, 2006, 142: 254-264.
pmid: 16844839 |
| [17] |
Forestan C, Meda S, Varotto S. ZmPIN1-mediated auxin transport is related to cellular differentiation during maize embryogenesis and endosperm development. Plant Physiol, 2010, 152: 1373-1390.
doi: 10.1104/pp.109.150193 pmid: 20044449 |
| [18] |
马娟, 曹言勇, 李会勇. 玉米穗轴粗全基因组关联分析. 作物学报, 2021, 47: 1228-1238.
doi: 10.3724/SP.J.1006.2021.03048 |
|
Ma J, Cao Y Y, Li H Y. Genome-wide association study of ear cob diameter in maize. Acta Agron Sin, 2021, 47: 1228-1238. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2021.03048 |
|
| [19] | 殷芳冰, 王成, 龙艳, 董振营, 万向元. 玉米雌穗性状遗传分析与形成机制. 中国生物工程杂志, 2021, 41(12): 30-46. |
| Yin F B, Wang C, Long Y, Dong Z Y, Wan X Y. Progress on dissecting genetic architecture and formation mechanism of maize ear traits. China Biotechnol, 2021, 41(12): 30-46. (in Chinese with English abstract) | |
| [20] |
Lu M, Xie C X, Li X H, Hao Z F, Li M S, Weng J F, Zhang D G, Bai L, Zhang S H. Mapping of quantitative trait loci for kernel row number in maize across seven environments. Mol Breed, 2011, 28: 143-152.
doi: 10.1007/s11032-010-9468-3 |
| [21] |
Karen S P, Lopes S C J, Pereira S A, Augusto F G A. QTL mapping for yield components in a tropical maize population using microsatellite markers. Hereditas, 2008, 145: 194-203.
doi: 10.1111/j.0018-0661.2008.02065.x |
| [22] |
Tian B H, Wang J H, Wang G Y. Confirmation of a major QTL on chromosome 10 for maize kernel row number in different environments. Plant Breed, 2014, 133: 184-188.
doi: 10.1111/pbr.12132 |
| [23] |
Liu L, Du Y F, Huo D A, Wang M, Shen X M, Yue B, Qiu F Z, Zheng Y L, Yan J B, Zhang Z X. Genetic architecture of maize kernel row number and whole genome prediction. Theor Appl Genet, 2015, 128: 2243-2254.
doi: 10.1007/s00122-015-2581-2 pmid: 26188589 |
| [24] |
Ma X Q, Tang J H, Teng W T, Yan J B, Meng Y J, Li J S. Epistatic interaction is an important genetic basis of grain yield and its components in maize. Mol Breed, 2007, 20: 41-51.
doi: 10.1007/s11032-006-9071-9 |
| [25] |
Yang C, Liu J, Rong T Z. Detection of quantitative trait loci for ear row number in F2 populations of maize. Genet Mol Res, 2015, 14: 14229-14238.
doi: 10.4238/2015.November.13.6 pmid: 26600480 |
| [26] |
Yan J B, Tang H, Huang Y Q, Zheng Y L, Li J S. Quantitative trait loci mapping and epistatic analysis for grain yield and yield components using molecular markers with an elite maize hybrid. Euphytica, 2006, 149: 121-131.
doi: 10.1007/s10681-005-9060-9 |
| [27] |
Brown P J, Upadyayula N, Mahone G S, Tian F, Bradbury P J, Myles S, Holland J B, Flint-Garcia S, McMullen M D, Buckler E S, Rocheford T R. Distinct genetic architectures for male and female inflorescence traits of maize. PLoS Genet, 2011, 7: e1002383.
doi: 10.1371/journal.pgen.1002383 |
| [28] |
Huo D A, Ning Q, Shen X M, Liu L, Zhang Z X. QTL mapping of kernel number-related traits and validation of one major QTL for ear length in maize. PLoS One, 2016, 11: e0155506.
doi: 10.1371/journal.pone.0155506 |
| [29] |
Yang N, Lu Y L, Yang X H, Huang J, Zhou Y, Ali F, Wen W W, Liu J, Li J S, Yan J B. Genome wide association studies using a new nonparametric model reveal the genetic architecture of 17 agronomic traits in an enlarged maize association panel. PLoS Genet, 2014, 10: e1004573.
doi: 10.1371/journal.pgen.1004573 |
| [30] |
Gallavotti A, Barazesh S, Malcomber S, Hall D, Jackson D, Schmidt R J, McSteen P. Sparse inflorescence1 encodes a monocot-specific YUCCA-like gene required for vegetative and reproductive development in maize. Proc Natl Acad Sci USA, 2008, 105: 15196-15201.
doi: 10.1073/pnas.0805596105 |
| [31] |
Phillips K A, Skirpan A L, Liu X, Christensen A, Slewinski T L, Hudson C, Barazesh S, Cohen J D, Malcomber S, McSteen P. Vanishing tassel2 encodes a grass-specific tryptophan aminotransferase required for vegetative and reproductive development in maize. Plant Cell, 2011, 23: 550-566.
doi: 10.1105/tpc.110.075267 |
| [32] |
Barazesh S, McSteen P. Barren inflorescence1 functions in organogenesis during vegetative and inflorescence development in maize. Genetics, 2008, 179: 389-401.
doi: 10.1534/genetics.107.084079 pmid: 18493061 |
| [33] |
McSteen P, Hake S. Barren inflorescence2 regulates axillary meristem development in the maize inflorescence. Development, 2001, 128: 2881-2891.
doi: 10.1242/dev.128.15.2881 pmid: 11532912 |
| [34] |
McSteen P, Malcomber S, Skirpan A, Lunde C, Wu X T, Kellogg E, Hake S. Barren inflorescence2 encodes a co-ortholog of the PINOID serine/threonine kinase and is required for organogenesis during inflorescence and vegetative development in maize. Plant Physiol, 2007, 144: 1000-1011.
doi: 10.1104/pp.107.098558 pmid: 17449648 |
| [35] |
Kato H, Nishihama R, Weijers D, Kohchi T. Evolution of nuclear auxin signaling: lessons from genetic studies with basal land plants. J Exp Bot, 2018, 69: 291-301.
doi: 10.1093/jxb/erx267 pmid: 28992186 |
| [36] |
Gallavotti A, Zhao Q, Kyozuka J, Meeley R B, Ritter M K, Doebley J F, Pè M E, Schmidt R J. The role of barren stalk1 in the architecture of maize. Nature, 2004, 432: 630-635.
doi: 10.1038/nature03148 |
| [37] |
Sigmon B, Vollbrecht E. Evidence of selection at the ramosa1 locus during maize domestication. Mol Ecol, 2010, 19: 1296-1311.
doi: 10.1111/j.1365-294X.2010.04562.x |
| [38] | 肖朝文, 傅永福. AT-hook蛋白的研究进展. 中国农业科技导报, 2009, 11(5): 12-16. |
| Xiao C W, Fu Y F. Research progress in AT-hook proteins. J Agric Sci Technol, 2009, 11(5): 12-16. (in Chinese with English abstract) | |
| [39] |
Gallavotti A, Malcomber S, Gaines C, Stanfield S, Whipple C, Kellogg E, Schmidt R J. BARREN STALK FASTIGIATE1 is an AT-hook protein required for the formation of maize ears. Plant Cell, 2011, 23: 1756-1771.
doi: 10.1105/tpc.111.084590 |
| [40] |
Jia H T, Li M F, Li W Y, Liu L, Jian Y N, Yang Z X, Shen X M, Ning Q, Du Y F, Zhao R, Jackson D, Yang X H, Zhang Z X. A serine/threonine protein kinase encoding gene KERNEL NUMBER PER ROW6 regulates maize grain yield. Nat Commun, 2020, 11: 988.
doi: 10.1038/s41467-020-14746-7 |
| [1] | 杨俊芳, 王宙, 乔麟轶, 王亚, 赵宜婷, 张宏斌, 申登高, 王宏伟, 曹越. 基于高密度遗传图谱的蓖麻种子大小性状QTL定位[J]. 作物学报, 2023, 49(3): 719-730. |
| [2] | 许加波, 吴鹏昊, 黄博文, 陈占辉, 马月虹, 任姣姣. 利用F2:3家系来源单倍体定位玉米雄穗相关性状QTL及全基因组选择[J]. 作物学报, 2023, 49(3): 622-633. |
| [3] | 马雅杰, 鲍建喜, 高悦欣, 李雅楠, 秦文萱, 王彦博, 龙艳, 李金萍, 董振营, 万向元. 玉米株高和穗位高性状全基因组关联分析[J]. 作物学报, 2023, 49(3): 647-661. |
| [4] | 刘月, 明博, 李姚姚, 王克如, 侯鹏, 薛军, 李少昆, 谢瑞芝. 基于根冠协调发展的东北春玉米高产种植密度分析[J]. 作物学报, 2023, 49(3): 795-807. |
| [5] | 刘姗姗, 庞婷, 袁晓婷, 罗凯, 陈平, 付智丹, 王小春, 杨峰, 雍太文, 杨文钰. 种间距对不同结瘤特性套作大豆根瘤生长及固氮潜力的影响[J]. 作物学报, 2023, 49(3): 833-844. |
| [6] | 方娅婷, 任涛, 张顺涛, 周橡棋, 赵剑, 廖世鹏, 丛日环, 鲁剑巍. 氮磷钾肥对旱地和水田油菜产量及养分利用的影响差异[J]. 作物学报, 2023, 49(3): 772-783. |
| [7] | 邓照, 蒋环琪, 程丽沙, 刘睿, 黄敏, 李曼菲, 杜何为. 利用WGCNA鉴定玉米非生物胁迫相关基因共表达网络[J]. 作物学报, 2023, 49(3): 672-686. |
| [8] | 宋杰, 王少祥, 李亮, 黄金苓, 赵斌, 张吉旺, 任佰朝, 刘鹏. 施钾量对夏玉米氮、磷、钾吸收利用和籽粒产量的影响[J]. 作物学报, 2023, 49(2): 539-551. |
| [9] | 刘梦, 张垚, 葛均筑, 周宝元, 吴锡冬, 杨永安, 侯海鹏. 不同降雨年型施氮量与收获期对夏玉米产量及氮肥利用效率的影响[J]. 作物学报, 2023, 49(2): 497-510. |
| [10] | 徐彤, 吕艳杰, 邵玺文, 耿艳秋, 王永军. 不同时期化控对密植玉米冠层结构及籽粒灌浆特性的影响[J]. 作物学报, 2023, 49(2): 472-484. |
| [11] | 杨硕, 武阳春, 刘鑫磊, 唐晓飞, 薛永国, 曹旦, 王婉, 刘亭萱, 祁航, 栾晓燕, 邱丽娟. 大豆蛋白含量主效位点qPRO-20-1的精细定位[J]. 作物学报, 2023, 49(2): 310-320. |
| [12] | 孙智超, 张吉旺. 弱光胁迫影响玉米产量形成的生理机制及调控效应[J]. 作物学报, 2023, 49(1): 12-23. |
| [13] | 徐凯, 郑兴飞, 张红燕, 胡中立, 宁子岚, 李兰芝. 基于NCII遗传交配设计的籼稻抽穗期全基因组关联分析[J]. 作物学报, 2023, 49(1): 86-96. |
| [14] | 陈冰洁, 张富粮, 杨硕, 李晓立, 何堂庆, 张晨曦, 田明慧, 吴梅, 郝晓峰, 张学林. 不同形态氮肥下丛枝菌根真菌对玉米灌浆期生理特性及产量和品质的影响[J]. 作物学报, 2023, 49(1): 249-261. |
| [15] | 张静, 王洪章, 任昊, 殷复伟, 吴红燕, 赵斌, 张吉旺, 任佰朝, 戴爱斌, 刘鹏. 夏玉米根系构型与抗根倒性能间的关系[J]. 作物学报, 2023, 49(1): 188-199. |
|
||