欢迎访问作物学报,今天是
作物学报 ›› 2019, Vol. 45 ›› Issue (11): 1638-1648.doi: 10.3724/SP.J.1006.2019.94003
刘浩,鲁清,李海芬,李少雄,陈小平,梁炫强( ),洪彦彬()
),洪彦彬()
LIU Hao,LU Qing,LI Hai-Fen,LI Shao-Xiong,CHEN Xiao-Ping,LIANG Xuan-Qiang(),HONG Yan-Bin()
摘要:
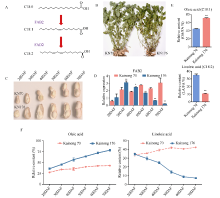
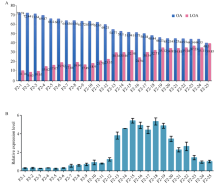
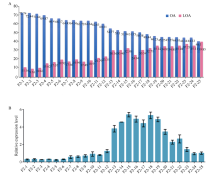
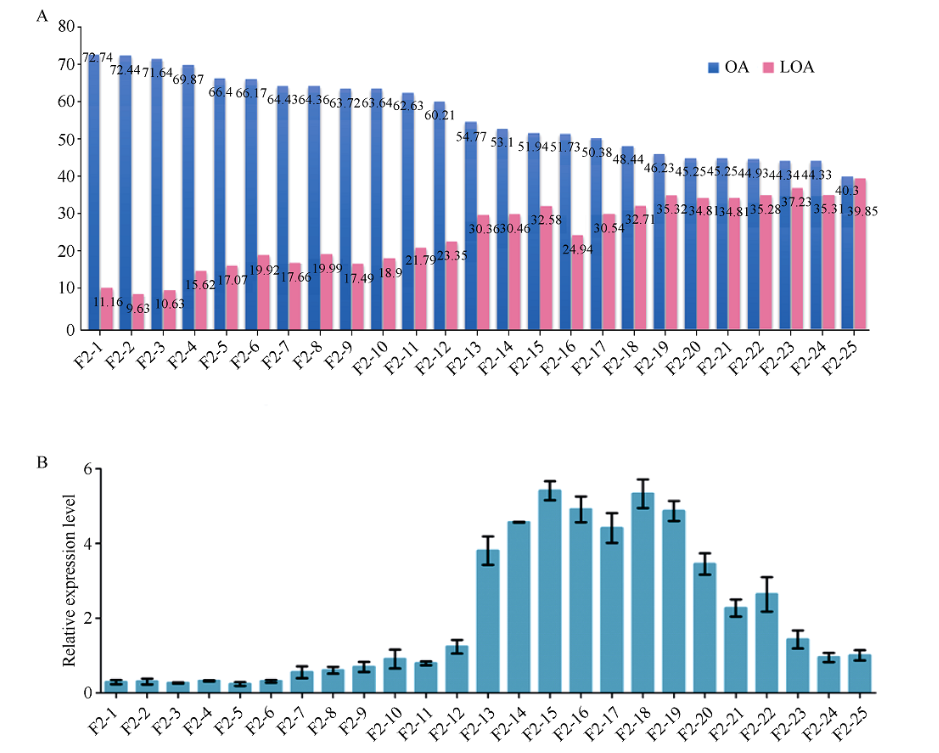
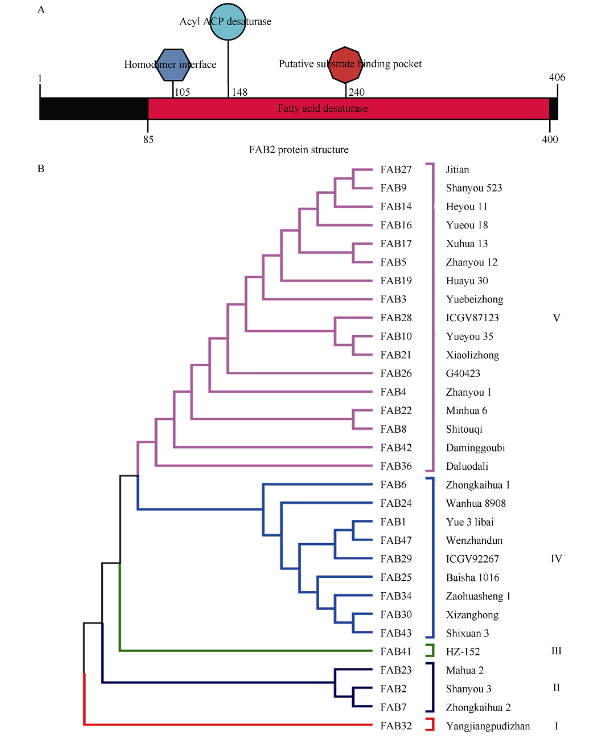
FAB2位于油酸合成通路的上游, 编码硬脂酰-ACP脱饱和酶, 调控硬脂酸(C18:0)向油酸(C18:1)转化。本研究发现高油酸品种开农176种子发育前期FAB2的表达量升高, 而成熟期油酸过量积累会抑制FAB2的表达。利用开农176与开农70构建F2杂交群体, 发现当植株油酸含量超过60%时会从整体水平上抑制FAB2的表达。种子发育前期, 油酸不断积累会导致过氧化物酶活性升高, 并且活性氧含量随之增加; 但是在种子发育后期均降低, 该结果与FAB2的表达量变化趋势相同。亚细胞定位结果表明, FAB2与FAD2分别定位于叶绿体与内质网。FAB2编码区序列多态性分析显示, 该蛋白序列氮端的氨基酸结构缺失可能会导致硬脂酸含量升高。FAB2启动子序列存在大量AT碱基的富集区域, 并且含有光响应、激素调控、转录因子结合的保守顺式元件。本研究发现过量积累的油酸会激活过氧化物酶介导的活性氧信号途径, 进而通过细胞核内的未知转录因子调节上游基因FAB2的表达量, 该结果不仅拓展了对FAB2的功能认知, 也为培育高油酸花生品种提供了相关的理论指导。
| [1] | 鲁清, 李少雄, 陈小平, 周桂元, 洪彦彬, 李海芬, 梁炫强 . 我国南方产区花生育种现状、存在问题及育种建议. 中国油料作物学报, 2017,39:556-566. |
| Lu Q, Li S X, Chen X P, Zhou G Y, Hong Y B, Li H F, Liang X Q . Current situation, problems and suggestions of peanut breeding in southern china. Chin J Oil Crop Sci, 2017,39:556-566 (in Chinese with English abstract). | |
| [2] | Gupta K, Kayam G, Faigenboim-Doron A, Clevenger J, Ozias-Akins P, Hovav R . Gene expression profiling during seed-filling process in peanut with emphasis on oil biosynthesis networks. Plant Sci, 2016,248:116-127. |
| [3] | Akhtar S, Khalid N, Ahmed I, Shahzad A, Suleria H A . Physicochemical characteristics, functional properties, and nutritional benefits of peanut oil: a review. Crit Rev Food Sci Nutr, 2014,54:1562-1575. |
| [4] | Isleib T G, Pattee H E, Sanders T H, Hendrix K W, Dean L O . Compositional and sensory comparisons between normal- and high-oleic peanuts. J Agric Food Chem, 2006,54:1759-1763. |
| [5] | Dar A A, Choudhury A R, Kancharla P K, Arumugam N . The FAD2 gene in plants: occurrence, regulation, and role. Front Plant Sci, 2017,8:1789. doi: 10.3389/fpls.2017.01789. |
| [6] | Patel M, Jung S, Moore K, Powell G, Ainsworth C, Abbott A . High-oleate peanut mutants result from a MITE insertion into the FAD2 gene. Theor Appl Genet, 2004,108:1492-1502. |
| [7] | 刘芳, 王积军, 汤松 . 我国高油酸花生品种选育与推广应用. 中国农技推广, 2017,33(1):14-15. |
| Liu F, Wang J J, Tang S . High-oleic peanut variety cultivation and popularized application in china. China Agric Technol Extension, 2017,33(1):14-15 (in Chinese with English abstract). | |
| [8] | Chi X, Yang Q, Pan L, Chen M, He Y, Yang Z, Yu S . Isolation and characterization of fatty acid desaturase genes from peanut (Arachis hypogaea L.). Plant Cell Rep, 2011,30:1393-1404. |
| [9] | Kachroo A, Shanklin J, Whittle E, Lapchyk L, Hildebrand D, Kachroo P . The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Mol Biol, 2007,63:257-271. |
| [10] | Mandal M K, Chandra-Shekara A C, Jeong R D, Yu K, Zhu S, Chanda B, Navarre D, Kachroo A, Kachroo P . Oleic acid- dependent modulation of NITRIC OXIDE ASSOCIATED1 protein levels regulates nitric oxide-mediated defense signaling in Arabidopsis. Plant Cell, 2012,24:1654-1674. |
| [11] | Shi J, Cao Y, Fan X R, Li M, Wang Y, Ming F . A rice microsomal delta-12 fatty acid desaturase can enhance resistance to cold stress in yeast and Oryza sativa. Mol Breed, 2012,29:743-757. |
| [12] | Jiang C J, Shimono M, Maeda S, Inoue H, Mori M, Hasegawa M, Sugano S, Takatsuji H . Suppression of the rice fatty-acid desaturase gene OsSSI2 enhances resistance to blast and leaf blight diseases in rice. Mol Plant Microbe Interact, 2009,22:820-829. |
| [13] | Song N, Hu Z, Li Y, Li C, Peng F, Yao Y, Peng H, Ni Z, Xie C, Sun Q . Overexpression of a wheat stearoyl-ACP desaturase (SACPD) gene TaSSI2 in Arabidopsis ssi2 mutant compromise its resistance to powdery mildew. Gene, 2013,524:220-227. |
| [14] | Chi X, Zhang Z, Chen N, Zhang X, Wang M, Chen M, Wang T, Pan L, Chen J, Yang Z, Guan X, Yu S . Isolation and functional analysis of fatty acid desaturase genes from peanut (Arachis hypogaea L.). PLoS One, 2017,12:e189759. |
| [15] | Liu H, Li H, Gu J, Deng L, Ren L, Hong Y, Lu Q, Chen X, Liang X . Identification of the candidate proteins related to oleic acid accumulation during peanut (Arachis hypogaea L.) seed development through comparative proteome analysis. Int J Mol Sci, 2018, 19 (4). pii: E1235. doi: 10.3390/ijms19041235. |
| [16] | Wen S, Liu H, Li X, Chen X, Hong Y, Li H, Lu Q, Liang X . TALEN-mediated targeted mutagenesis of fatty acid desaturase 2 (FAD2) in peanut(Arachis hypogaea L.) promotes the accumulation of oleic acid. Plant Mol Biol, 2018,97:177-185. |
| [17] | Sui N, Wang Y, Liu S, Yang Z, Wang F, Wan S . Transcriptomic and physiological evidence for the relationship between unsaturated fatty acid and salt stress in peanut. Front Plant Sci, 2018,9:7. doi: 10.3389/fpls.2018.00007. |
| [18] | Zhao J, Favero D S, Qiu J, Roalson E H, Neff M M . Insights into the evolution and diversification of the AT-hook motif nuclear localized gene family in land plants. BMC Plant Biol, 2014,14:266. doi: 10.1186/s12870-014-0266-7. |
| [19] | Chen X, Zhu W, Azam S, Li H, Zhu F, Li H, Hong Y, Liu H, Zhang E, Wu H, Yu S, Zhou G, Li S, Zhong N, Wen S, Li X, Knapp S J, Ozias-Akins P, Varshney R K, Liang X . Deep sequencing analysis of the transcriptomes of peanut aerial and subterranean young pods identifies candidate genes related to early embryo abortion. Plant Biotechnol J, 2013,11:115-127. |
| [20] | Grevengoed T J, Klett E L, Coleman R A . Acyl-CoA metabolism and partitioning. Annu Rev Nutr, 2014,34:1-30. |
| [21] | Yang W, Dong R, Liu L, Hu Z, Li J, Wang Y, Ding X, Chu Z . A novel mutant allele of SSI2 confers a better balance between disease resistance and plant growth inhibition on Arabidopsis thaliana. BMC Plant Biol, 2016,16:208. doi: 10.1186/s12870-016-0898-x. |
| [22] | Theodoulou F L, Eastmond P J . Seed storage oil catabolism: a story of give and take. Curr Opin Plant Biol, 2012,15:322-328. |
| [23] | Kao Y T, Gonzalez K L, Bartel B . Peroxisome function, biogenesis, and dynamics in plants. Plant Physiol, 2018,176:162-177. |
| [24] | Wasternack C, Song S . Jasmonates: biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J Exp Bot, 2017,68:1303-1321. |
| [1] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [2] | 李海芬, 魏浩, 温世杰, 鲁清, 刘浩, 李少雄, 洪彦彬, 陈小平, 梁炫强. 花生电压依赖性阴离子通道基因(AhVDAC)的克隆及在果针向地性反应中表达分析[J]. 作物学报, 2022, 48(6): 1558-1565. |
| [3] | 刘嘉欣, 兰玉, 徐倩玉, 李红叶, 周新宇, 赵璇, 甘毅, 刘宏波, 郑月萍, 詹仪花, 张刚, 郑志富. 耐三唑并嘧啶类除草剂花生种质创制与鉴定[J]. 作物学报, 2022, 48(4): 1027-1034. |
| [4] | 丁红, 徐扬, 张冠初, 秦斐斐, 戴良香, 张智猛. 不同生育期干旱与氮肥施用对花生氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 695-703. |
| [5] | 黄莉, 陈玉宁, 罗怀勇, 周小静, 刘念, 陈伟刚, 雷永, 廖伯寿, 姜慧芳. 花生种子大小相关性状QTL定位研究进展[J]. 作物学报, 2022, 48(2): 280-291. |
| [6] | 荐红举, 尚丽娜, 金中辉, 丁艺, 李燕, 王季春, 胡柏耿, Vadim Khassanov, 吕典秋. 马铃薯PIF家族成员鉴定及其对高温胁迫的响应分析[J]. 作物学报, 2022, 48(1): 86-98. |
| [7] | 汪颖, 高芳, 刘兆新, 赵继浩, 赖华江, 潘小怡, 毕晨, 李向东, 杨东清. 利用WGCNA鉴定花生主茎生长基因共表达模块[J]. 作物学报, 2021, 47(9): 1639-1653. |
| [8] | 王建国, 张佳蕾, 郭峰, 唐朝辉, 杨莎, 彭振英, 孟静静, 崔利, 李新国, 万书波. 钙与氮肥互作对花生干物质和氮素积累分配及产量的影响[J]. 作物学报, 2021, 47(9): 1666-1679. |
| [9] | 石磊, 苗利娟, 黄冰艳, 高伟, 张忠信, 齐飞艳, 刘娟, 董文召, 张新友. 花生AhFAD2-1基因启动子及5'-UTR内含子功能验证及其低温胁迫应答[J]. 作物学报, 2021, 47(9): 1703-1711. |
| [10] | 高芳, 刘兆新, 赵继浩, 汪颖, 潘小怡, 赖华江, 李向东, 杨东清. 北方主栽花生品种的源库特征及其分类[J]. 作物学报, 2021, 47(9): 1712-1723. |
| [11] | 张鹤, 蒋春姬, 殷冬梅, 董佳乐, 任婧瑶, 赵新华, 钟超, 王晓光, 于海秋. 花生耐冷综合评价体系构建及耐冷种质筛选[J]. 作物学报, 2021, 47(9): 1753-1767. |
| [12] | 薛晓梦, 吴洁, 王欣, 白冬梅, 胡美玲, 晏立英, 陈玉宁, 康彦平, 王志慧, 淮东欣, 雷永, 廖伯寿. 低温胁迫对普通和高油酸花生种子萌发的影响[J]. 作物学报, 2021, 47(9): 1768-1778. |
| [13] | 郝西, 崔亚男, 张俊, 刘娟, 臧秀旺, 高伟, 刘兵, 董文召, 汤丰收. 过氧化氢浸种对花生种子发芽及生理代谢的影响[J]. 作物学报, 2021, 47(9): 1834-1840. |
| [14] | 张旺, 冼俊霖, 孙超, 王春明, 石丽, 于为常. CRISPR/Cas9编辑花生FAD2基因研究[J]. 作物学报, 2021, 47(8): 1481-1490. |
| [15] | 戴良香, 徐扬, 张冠初, 史晓龙, 秦斐斐, 丁红, 张智猛. 花生根际土壤细菌群落多样性对盐胁迫的响应[J]. 作物学报, 2021, 47(8): 1581-1592. |
|
||