欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (10): 1903-1912.doi: 10.3724/SP.J.1006.2021.03060
周练( ), 刘朝显, 陈秋栏, 王文琴, 姚顺, 赵子堃, 朱思颖, 洪祥德, 熊雨涵, 蔡一林*()
), 刘朝显, 陈秋栏, 王文琴, 姚顺, 赵子堃, 朱思颖, 洪祥德, 熊雨涵, 蔡一林*()
ZHOU Lian(), LIU Chao-Xian, CHEN Qiu-Lan, WANG Wen-Qin, YAO Shun, ZHAO Zi-Kun, ZHU Si-Ying, HONG Xiang-De, XIONG Yu-Han, CAI Yi-Lin*()
摘要:
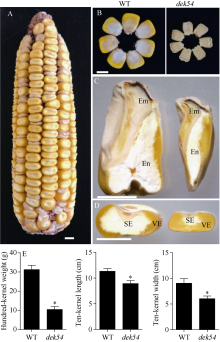
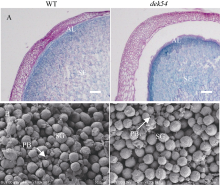
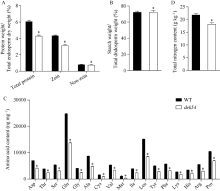
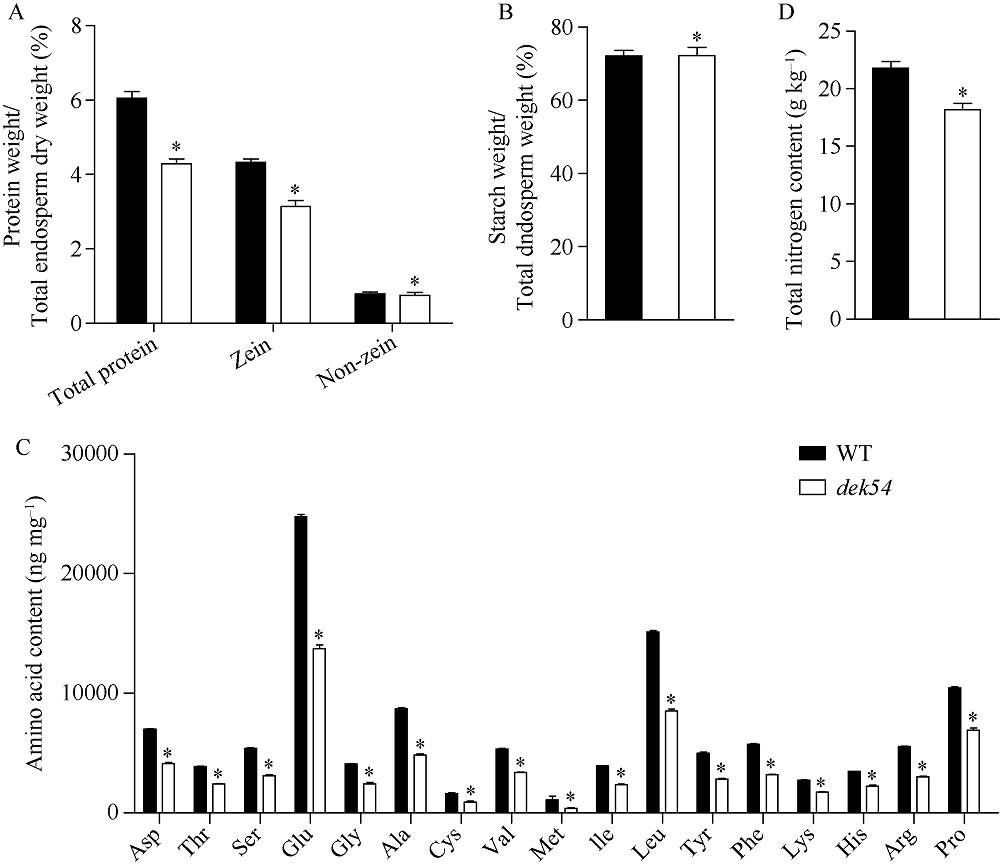
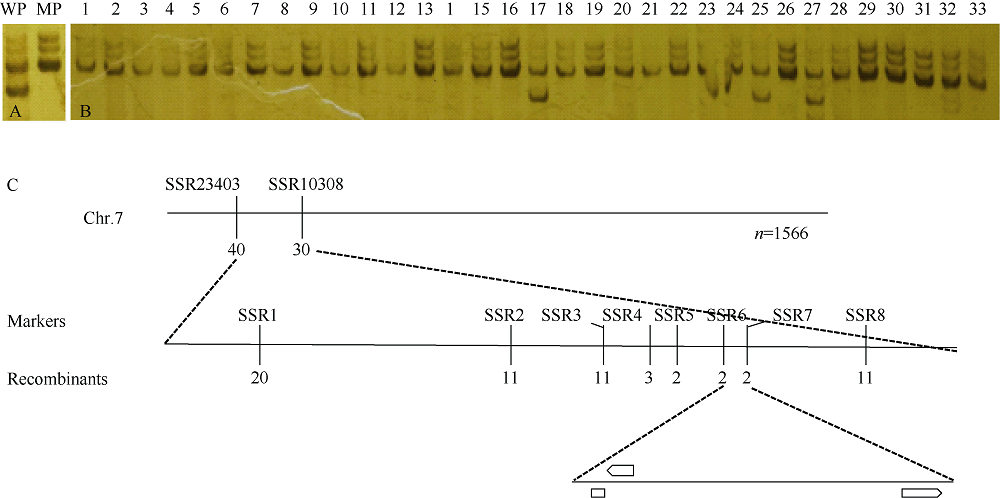
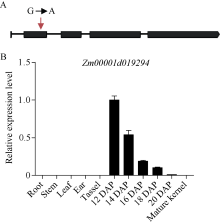
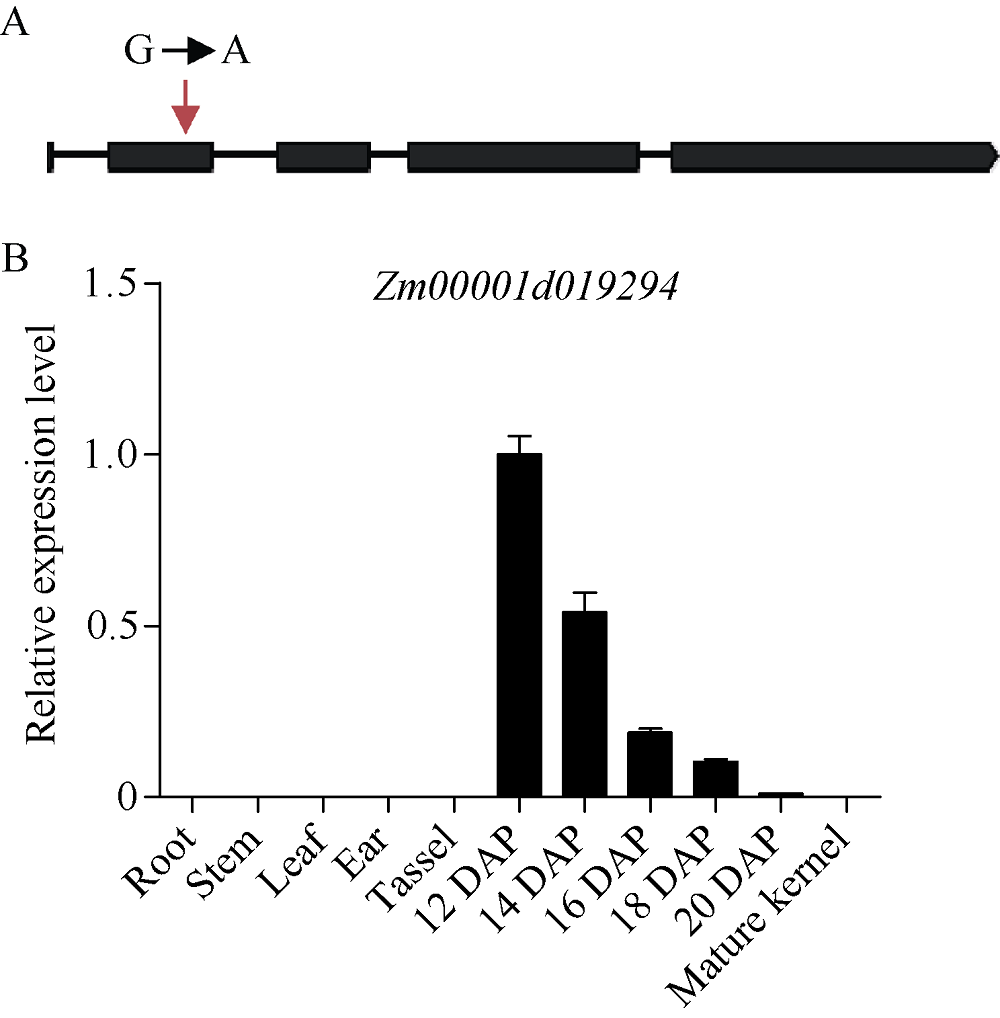
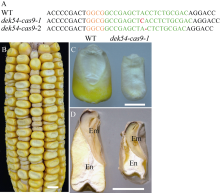
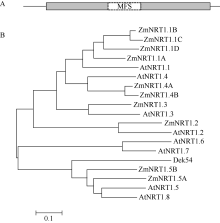
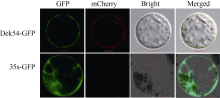
玉米籽粒与产量和营养品质密切相关, 控制籽粒发育基因的功能研究对解析籽粒发育分子机制, 提高玉米产量, 改善籽粒营养品质提供重要依据。利用甲基磺酸乙酯(ethyl methanesulfonate, EMS)处理B73花粉, 筛选到一个玉米籽粒缺陷突变体defective kernel 54 (dek54)。dek54表现出成熟籽粒变小、皱缩、颜色发白等特征; 遗传分析表明dek54是一个单基因控制的隐性突变体。石蜡切片显示dek54淀粉胚乳细胞形状不规则且排列致密, 扫描电镜观察成熟籽粒胚乳中心区域发现dek54淀粉粒周围蛋白体比野生型少且排列疏松。dek54成熟籽粒的总蛋白、醇溶蛋白、各氨基酸组分和全氮含量相比野生型都显著降低。利用F2分离群体中的1566个dek54单株, 把dek54定位在7号染色体标记SSR6和SSR7之间, 物理位置约为290 kb。该区间有3个基因, 基因测序发现Zm00001d019294基因第2个外显子上第351个碱基由G突变为A, 从而导致蛋白翻译的提前终止。该基因在玉米籽粒中特异性表达, 且在12 DAP (days after pollination)籽粒中表达量最高。通过CRISPR/Cas9系统进行靶向突变确定候选基因Zm00001d019294导致该突变表型。Dek54编码一个与ZmNRT1.5 (nitrate transporter)具有较高同源性的MFS (major facilitator superfamily)家族蛋白并定位在玉米原生质体的细胞质膜。该研究为揭示dek54在玉米籽粒发育的分子机制奠定了重要基础。
| [1] |
Sabelli P A, Larkins B A. The development of endosperm in grasses. Plant Physiol, 2009, 149:14-26.
doi: 10.1104/pp.108.129437 pmid: 19126691 |
| [2] |
Olsen O A. Endosperm development: cellularization and cell fate specification. Annu Rev Plant Physiol Plant Mol Biol, 2001, 52:233-267.
doi: 10.1146/annurev.arplant.52.1.233 |
| [3] |
Lid S E, Gruis D, Jung R, Lorentzen J A, Ananiev E, Chamberlin M, Niu X, Meeley R, Nichols S, Olsen O A. The defective kernel 1 (dek1) gene required for aleurone cell development in the endosperm of maize grains encodes a membrane protein of the calpain gene superfamily. Proc Natl Acad Sci USA, 2002, 99:5460-5465.
doi: 10.1073/pnas.042098799 |
| [4] |
Demko V, Perroud P F, Johansen W, Delwiche C F, Cooper E D, Remme P, Ako A E, Kugler K G, Mayer K F, Quatrano R, Olsen O A. Genetic analysis of DEFECTIVE KERNEL1 loop function in three-dimensional body patterning in Physcomitrella patens. Plant Physiol, 2014, 166:903-919.
doi: 10.1104/pp.114.243758 |
| [5] |
Becraft P W, Li K, Dey N, Asuncion-Crabb Y. The maize dek1 gene functions in embryonic pattern formation and cell fate specification. Development, 2002, 129:5217-5225.
pmid: 12399313 |
| [6] |
Tian Q, Olsen L, Sun B, Lid S E, Brown R C, Lemmon B E, Fosnes K, Gruis D F, Opsahl-Sorteberg H G, Otegui M S, Olsen O A. Subcellular localization and functional domain studies of DEFECTIVE KERNEL1 in maize and Arabidopsis suggest a model for aleurone cell fate specification involving CRINKLY4 and SUPERNUMERARY ALEURONE LAYER1. Plant Cell, 2007, 19:3127-3145.
doi: 10.1105/tpc.106.048868 |
| [7] |
Qi W, Yang Y, Feng X, Zhang M, Song R. Mitochondrial function and maize kernel development requires Dek2, a pentatricopeptide repeat Protein involved in nad1 mRNA splicing. Genetics, 2017, 205:239-249.
doi: 10.1534/genetics.116.196105 |
| [8] |
He Y, Wang J, Qi W, Song R. Maize Dek15 encodes the cohesin-loading complex subunit SCC4 and is essential for chromosome segregation and kernel development. Plant Cell, 2019, 31:465-485.
doi: 10.1105/tpc.18.00921 |
| [9] |
Wang G, Zhong M, Shuai B, Song J, Zhang J, Han L, Ling H, Tang Y, Wang G, Song R. E+ subgroup PPR protein defective kernel 36 is required for multiple mitochondrial transcripts editing and seed development in maize and Arabidopsis. New Phytol, 2017, 214:1563-1578.
doi: 10.1111/nph.2017.214.issue-4 |
| [10] |
Dai D, Luan S, Chen X, Wang Q, Feng Y, Zhu C, Qi W, Song R. Maize Dek37 encodes a P-type PPR protein that affects cis-splicing of mitochondrial nad2 intron 1 and seed development. Genetics, 2018, 208:1069-1082.
doi: 10.1534/genetics.117.300602 |
| [11] |
Li X, Gu W, Sun S, Chen Z, Chen J, Song W, Zhao H, Lai J. Defective Kernel 39 encodes a PPR protein required for seed development in maize. J Integr Plant Biol, 2018, 60:45-64.
doi: 10.1111/jipb.v60.1 |
| [12] |
Qi W, Lu L, Huang S, Song R. Maize Dek44 encodes mitochondrial ribosomal protein L9 and is required for seed development. Plant Physiol, 2019, 180:2106-2119.
doi: 10.1104/pp.19.00546 |
| [13] |
Wang G, Wang F, Wang G, Wang F, Zhang X, Zhong M, Zhang J, Lin D, Tang Y, Xu Z, Song R. Opaque1 encodes a myosin XI motor protein that is required for endoplasmic reticulum motility and protein body formation in maize endosperm. Plant Cell, 2012, 24:3447-3462.
doi: 10.1105/tpc.112.101360 |
| [14] |
Schmidt R J, Burr F A, Burr B. Transposon tagging and molecular analysis of the maize regulatory locus opaque-2. Science, 1987, 238:960-963.
pmid: 2823388 |
| [15] |
Schmidt R J, Burr F A, Aukerman M J, Burr B. Maize regulatory gene opaque-2 encodes a protein with a “leucine-zipper” motif that binds to zein DNA. Proc Natl Acad Sci USA, 1990, 87:46-50.
doi: 10.1073/pnas.87.1.46 |
| [16] |
Myers A M, James M G, Lin Q, Yi G, Stinard P S, Hennen-Bierwagen T A, Becraft P W. Maize opaque5 encodes monogalactosyldiacylglycerol synthase and specifically affects galactolipids necessary for amyloplast and chloroplast function. Plant Cell, 2011, 23:2331-2347.
doi: 10.1105/tpc.111.087205 |
| [17] |
Wang G, Sun X, Wang G, Wang F, Gao Q, Sun X, Tang Y, Chang C, Lai J, Zhu L, Xu Z, Song R. Opaque7 encodes an acyl-activating enzyme-like protein that affects storage protein synthesis in maize endosperm. Genetics, 2011, 189:1281-1295.
doi: 10.1534/genetics.111.133967 |
| [18] |
Feng F, Qi W, Lü Y, Yan S, Xu L, Yang W, Yuan Y, Chen Y, Zhao H, Song R. OPAQUE11 is a central hub of the regulatory network for maize endosperm development and nutrient metabolism. Plant Cell, 2018, 30:375-396.
doi: 10.1105/tpc.17.00616 |
| [19] |
Sarika K, Hossain F, Muthusamy V, Zunjare R U, Baveja A, Goswami R, Thirunavukkarasu N, Jha S K, Gupta H S. Opaque16, a high lysine and tryptophan mutant, does not influence the key physico-biochemical characteristics in maize kernel. PLoS One, 2018, 13:e0190945.
doi: 10.1371/journal.pone.0190945 |
| [20] |
Kim C S, Gibbon B C, Gillikin J W, Larkins B A, Boston R S, Jung R. The maize Mucronate mutation is a deletion in the 16-kDa gamma-zein gene that induces the unfolded protein response. Plant J, 2006, 48:440-451.
doi: 10.1111/tpj.2006.48.issue-3 |
| [21] |
Kim C S, Hunter B G, Kraft J, Boston R S, Yans S, Jung R, Larkins B A. A defective signal peptide in a 19-kD alpha-zein protein causes the unfolded protein response and an opaque endosperm phenotype in the maize De*-B30 mutant. Plant Physiol, 2004, 134:380-387.
doi: 10.1104/pp.103.031310 |
| [22] |
Holding D R, Otegui M S, Li B, Meeley R B, Dam T, Hunter B G, Jung R, Larkins B A. The maize floury1 gene encodes a novel endoplasmic reticulum protein involved in zein protein body formation. Plant Cell, 2007, 19:2569-2582.
pmid: 17693529 |
| [23] |
Coleman C E, Lopes M A, Gillikin J W, Boston R S, Larkins B A. A defective signal peptide in the maize high-lysine mutant floury 2. Proc Natl Acad Sci USA, 1995, 92:6828-6831.
doi: 10.1073/pnas.92.15.6828 |
| [24] | 李强, 万建民. SSRHunter: 一个本地化的SSR位点搜索软件的开发. 遗传, 2005, 27:808-810. |
| Li Q, Wan J M. SSRHunter: development of a local searching software for SSR sites. Hereditas, 2005, 27:808-810. | |
| [25] |
Zhou L, Zhou J, Xiong Y, Liu C, Wang J, Wang G, Cai Y. Overexpression of a maize plasma membrane intrinsic protein ZmPIP1;1 confers drought and salt tolerance in Arabidopsis. PLoS One, 2018, 13:e0198639.
doi: 10.1371/journal.pone.0198639 |
| [26] |
Sheridan W F, Neuffer M G. Defective kernel mutants of maize: II. morphological and embryo culture studies. Genetics, 1980, 95:945-960.
pmid: 17249054 |
| [27] |
Zhu C, Jin G, Fang P, Zhang Y, Feng X, Tang Y, Qi W, Song R. Maize pentatricopeptide repeat protein DEK41 affects cis-splicing of mitochondrial nad4 intron 3 and is required for normal seed development. J Exp Bot, 2019, 70:3795-3808.
doi: 10.1093/jxb/erz193 |
| [28] |
Ren R C, Wang L L, Zhang L, Zhao Y J, Wu J W, Wei Y M, Zhang X S, Zhao X Y. DEK43 is a P-type pentatricopeptide repeat (PPR) protein responsible for the cis-splicing of nad4 in maize mitochondria. J Integr Plant Biol, 2020, 62:299-313.
doi: 10.1111/jipb.v62.3 |
| [29] | Dai D, Jin L, Huo Z, Yan S, Ma Z, Qi W, Song R. Pentatricopeptide repeat protein DEK46 is required for multi-sites mitochondrial RNA editing and maize seed development. J Exp Bot, 2020 |
| [30] |
Fujii S, Small I. The evolution of RNA editing and pentatricopeptide repeat genes. New Phytol, 2011, 191:37-47.
doi: 10.1111/j.1469-8137.2011.03746.x pmid: 21557747 |
| [31] |
Liang Z, Demko V, Wilson R C, Johnson K A, Ahmad R, Perroud P F, Quatrano R, Zhao S, Shalchian-Tabrizi K, Otegui M S, Olsen O A, Johansen W. The catalytic domain CysPc of the DEK1 calpain is functionally conserved in land plants. Plant J, 2013, 75:742-754.
doi: 10.1111/tpj.2013.75.issue-5 |
| [32] |
Saier M H, Jr Reddy V S, Tamang D G, Västermark A. The transporter classification database. Nucleic Acids Res, 2014, 42:D251-258.
doi: 10.1093/nar/gkt1097 |
| [33] |
Yan N. Structural biology of the Major Facilitator Superfamily transporters. Annu Rev Biophys, 2015, 44:257-283.
doi: 10.1146/annurev-biophys-060414-033901 |
| [34] |
Parker J L, Newstead S. Molecular basis of nitrate uptake by the plant nitrate transporter NRT1.1. Nature, 2014, 507:68-72.
doi: 10.1038/nature13116 |
| [35] |
Liu K H, Tsay Y F. Switching between the two action modes of the dual-affinity nitrate transporter CHL1 by phosphorylation. EMBO J, 2003, 22:1005-1013.
doi: 10.1093/emboj/cdg118 |
| [36] |
Liu K H, Huang C Y, Tsay Y F. CHL1 is a dual-affinity nitrate transporter of Arabidopsis involved in multiple phases of nitrate uptake. Plant Cell, 1999, 11:865-874.
pmid: 10330471 |
| [37] |
Lin S H, Kuo H F, Canivenc G, Lin C S, Lepetit M, Hsu P K, Tillard P, Lin H L, Wang Y Y, Tsai C B, Gojon A, Tsay Y F. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell, 2008, 20:2514-2528.
doi: 10.1105/tpc.108.060244 |
| [38] |
Li J Y, Fu Y L, Pike S M, Bao J, Tian W, Zhang Y, Chen C Z, Zhang Y, Li H M, Huang J, Li L G, Schroeder J I, Gassmann W, Gong J M. The Arabidopsis nitrate transporter NRT1.8 functions in nitrate removal from the xylem sap and mediates cadmium tolerance. Plant Cell, 2010, 22:1633-1646.
doi: 10.1105/tpc.110.075242 |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [4] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [5] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [6] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [7] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [8] | 王好让, 张勇, 于春淼, 董全中, 李微微, 胡凯凤, 张明明, 薛红, 杨梦平, 宋继玲, 王磊, 杨兴勇, 邱丽娟. 大豆突变体ygl2黄绿叶基因的精细定位[J]. 作物学报, 2022, 48(4): 791-800. |
| [9] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [10] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [11] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [12] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [13] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [14] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [15] | 张倩, 韩本高, 张博, 盛开, 李岚涛, 王宜伦. 控失尿素减施及不同配比对夏玉米产量及氮肥效率的影响[J]. 作物学报, 2022, 48(1): 180-192. |
|
||