欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (7): 1645-1657.doi: 10.3724/SP.J.1006.2022.14107
韩尚玲1( ), 霍轶琼1,2, 李辉1, 韩华蕊1, 侯思宇1,2,3, 孙朝霞1,2,3, 韩渊怀1,2,3, 李红英1,2,3,*()
), 霍轶琼1,2, 李辉1, 韩华蕊1, 侯思宇1,2,3, 孙朝霞1,2,3, 韩渊怀1,2,3, 李红英1,2,3,*()
HAN Shang-Ling1(), HUO Yi-Qiong1,2, LI Hui1, HAN Hua-Rui1, HOU Si-Yu1,2,3, SUN Zhao-Xia1,2,3, HAN Yuan-Huai1,2,3, LI Hong-Ying1,2,3,*()
摘要:
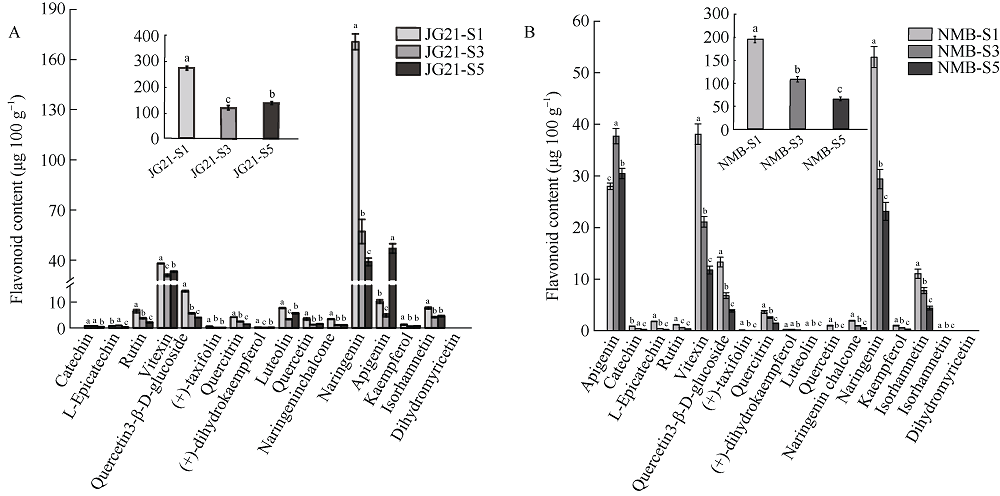
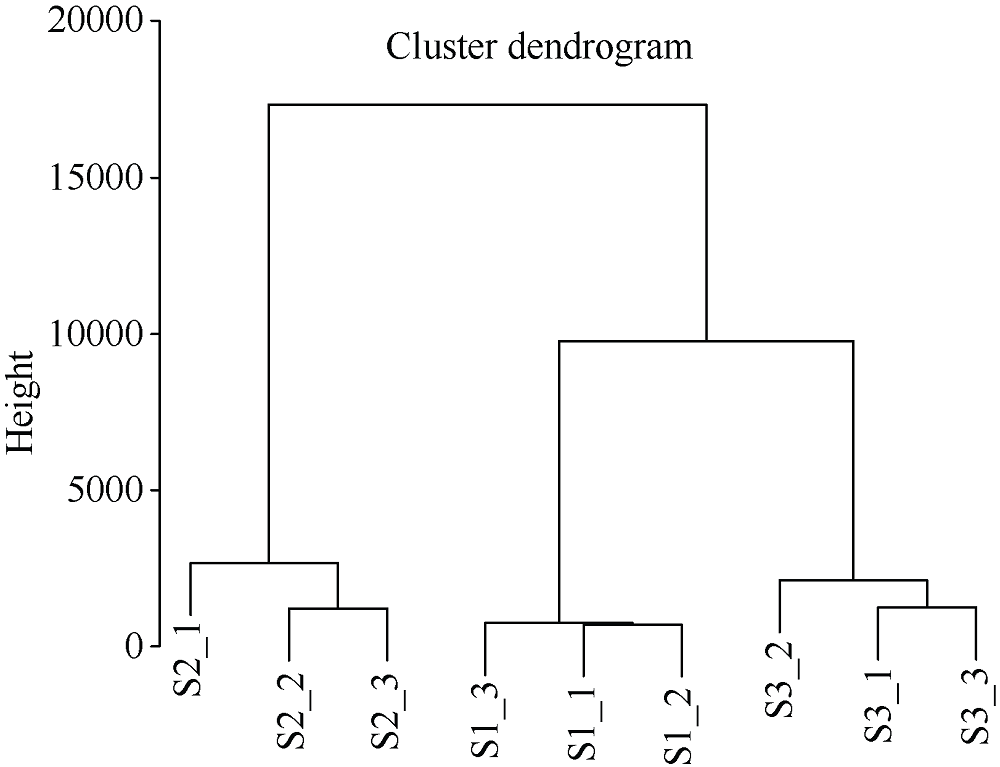
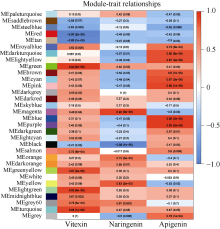
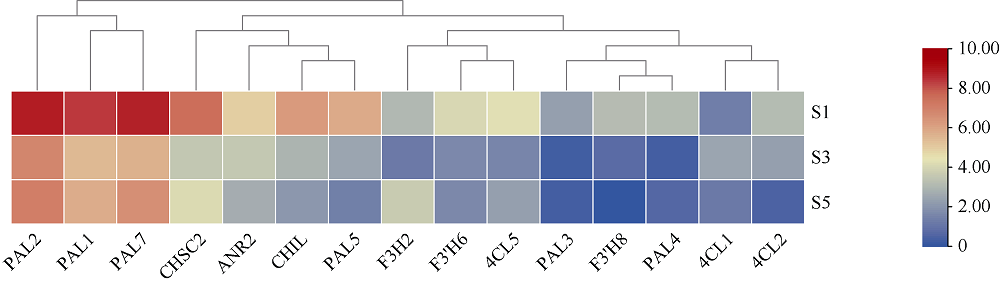
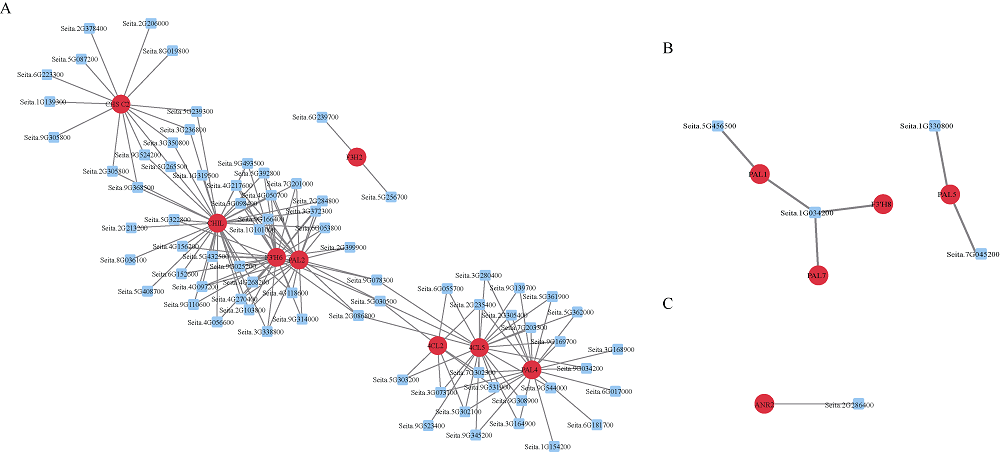
类黄酮是植物重要的次生代谢物质, 在植物生长发育中发挥着重要的作用, 此外, 类黄酮具有抗氧化活性, 对人体十分有益。谷子籽粒营养丰富全面, 是一种保健型杂粮, 深受我国人民喜爱。谷子作为C4模式植物, 正在受到越来越多的关注。目前谷子籽粒类黄酮代谢及调控机制研究较少。本研究利用高类黄酮品种晋谷21 (JG21)及低类黄酮品种牛毛白(NMB)谷穗为材料, 分析JG21与NMB小穗发育不同阶段类黄酮靶向代谢组及JG21小穗发育不同阶段转录组, 结合加权基因共表达网络分析挖掘可能参与调控类黄酮代谢的转录因子, 并在不同水平的类黄酮品种中进行表达分析初步验证。结果发现, 2个品种小穗中主要富集的类黄酮组分为芹菜素、牡荆素及柚皮素, 三者共占总类黄酮含量的79%以上。类黄酮代谢基因共表达网络中包含38,921个基因, 共划分为32个模块, 其中turquoise模块、green模块及magenta模块与类黄酮代谢显著相关。利用类黄酮代谢通路差异表达基因作为关键基因筛选出与类黄酮代谢调控相关的27个转录因子家族, 并通过启动子结合基序分析筛选获得11个转录因子。Pearson相关性分析表明, 11个转录因子中有7个候选转录因子可能参与类黄酮代谢, 分别为WRKY38、MYB4a、PI、WRKY15、WRKY62、MYB46、WRKY23。以上结果为研究谷子类黄酮代谢通路转录调控机制提供了新的候选基因, 为深入揭示类黄酮代谢调控机制奠定基础。
| [1] |
Williams C A, Grayer R J. Anthocyanins and other flavonoids. Nat Prod Rep, 2004, 21: 539-573.
doi: 10.1039/b311404j |
| [2] |
Buer C S, Imin N, Djordjevic M A. Flavonoids: new roles for old molecules. J Integr Plant Biol, 2010, 52: 98-111.
doi: 10.1111/j.1744-7909.2010.00905.x |
| [3] | Kozłowska A, Szostak-Wegierek D. Flavonoids-food sources and health benefits. Rocz Panstw Zakl Hig, 2014, 65: 79-85. |
| [4] |
Nakabayashi R, Yonekura-Sakakibara K, Urano K, Suzuki M, Yamada Y, Nishizawa T, Matsuda F, Kojima M, Sakakibara H, Shinozaki K, Michael A J, Tohge T, Yamazaki M, Saito K. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J, 2014, 77: 367-379.
doi: 10.1111/tpj.12388 |
| [5] |
Gouot J C, Smith J P, Holzapfel B P, Walker A R, Barril C. Grape berry flavonoids: a review of their biochemical responses to high and extreme high temperatures. J Exp Bot, 2019, 70: 397-423.
doi: 10.1093/jxb/ery392 |
| [6] | Pan J Q, Tong X R, Guo B L. Progress of effects of light on plant flavonoids. China J Chin Mater Med, 2016, 41: 3897-3903. |
| [7] |
Xu W J, Dubos C, Lepiniec L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci, 2015, 20: 176-185.
doi: 10.1016/j.tplants.2014.12.001 |
| [8] | 刁现民.谷子种质资源的深度分析和研究利用. 见: 2017年中国作物学会学术年会摘要集, 保定: 中国作物学会, 2017. p 1. |
| Diao X M. In-depth analysis and research utilization of foxtail millet germplasm resources. In: Abstract ppub of the Academic Annual Meeting of Chinese Crop Society in 2017Baoding: The Crop Science Society of China, 2017. p 1. (in Chinese) | |
| [9] | 徐玖亮, 温馨, 刁现民, 张福锁, 李学贤. 我国主要谷类杂粮的营养价值及保健功能. 粮食与饲料工业, 2021, (1): 27-35. |
| Xu J L, Wen X, Diao X M, Zhang F S, Li X X. Nutrition values and health effects of coarse cereals in China. Cereal Feed Ind, 2021, (1): 27-35. (in Chinese with English abstract) | |
| [10] |
Zhang Y K, Gao J H, Qie Q R, Yang Y L, Hou S Y, Wang X C, Li X K, Han Y H. Comparative analysis of flavonoid metabolites in foxtail millet (Setaria italica) with different eating quality. Life (Basel), 2021, 11: 578.
doi: 10.3390/life11060578 |
| [11] | 鲜小华, 王嘉, 徐新福, 曲存民, 卢坤, 李加纳, 刘列钊. 整合GWAS和WGCNA分析挖掘甘蓝型油菜黄籽微效作用位点. 作物学报, 2018, 44: 1105-1113. |
|
Xian X H, Wang J, Xu X F, Qu C M, Lu K, Li J N, Liu L Z. Mining yellow-seeded micro effect loci in B. napus by integrated GWAS and WGCNA analysis. Acta Agron Sin, 2018, 44: 1105-1113. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2018.01105 |
|
| [12] | 程旭.基于转录组和共表达网络分析的玫瑰类黄酮和萜类生物合成相关基因研究. 华中农业大学硕士学位论文,湖北武汉, 2016. |
| Cheng X. Research of Flavonoids and Terpenoids Biosynthesis Genes Based on Transcriptome and Co-expression Network Analysis. MS Thesis of Huazhong Agricultural University, Wuhan, Hubei, China, 2016. (in Chinese with English abstract) | |
| [13] |
Yang Z R, Zhang H S, Li X K, Shen H M, Gao J H, Hou S Y, Zhang B, Mayes S, Bennett M, Ma J X, Wu C Y, Sui Y, Han Y H, Wang X C. A mini foxtail millet with an Arabidopsis-like life cycle as a C4 model system. Nat Plants, 2020, 6: 1167-1178.
doi: 10.1038/s41477-020-0747-7 |
| [14] | Andrews S. Babraham bioinformatics-FastQC a quality control tool for high throughput sequence data. Bioinformatics, 2010, 26: 774-798. |
| [15] |
Bolger A M, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics, 2014, 30: 2114-2120.
doi: 10.1093/bioinformatics/btu170 |
| [16] |
Kim D, Langmead B, Salzberg S L. HISAT: a fast spliced aligner with low memory requirements. Nat Methods, 2015, 12: 357-360.
doi: 10.1038/NMETH.3317 |
| [17] |
Pertea M, Pertea G M, Antonescu C M, Chang T C, Mendell J T, Salzberg S L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat Biotechnol, 2015, 33: 290-295.
doi: 10.1038/nbt.3122 |
| [18] |
Love M I, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol, 2014, 15: 550.
doi: 10.1186/s13059-014-0550-8 |
| [19] | Fornes O, Castro-Mondragon J A, Khan A, van der Lee R, Zhang X, Richmond P A, Modi B P, Correard S, Gheorghe M, Baranašić D, Santana-Garcia W, Tan G, Chèneby J, Ballester B, Parcy F, Sandelin A, Lenhard B, Wasserman W W, Mathelier A. JASPAR 2020: update of the open-access database of transcription factor binding profiles. Nucleic Acids Res, 2020, 48: D87-D92. |
| [20] |
Monniaux M, McKim S M, Cartolano M, Thévenon E, Parcy F, Tsiantis M, Hay A. Conservation vs divergence in LEAFY and APETALA1 functions between Arabidopsis thaliana and Cardamine hirsuta. New Phytol, 2017, 216: 549-561.
doi: 10.1111/nph.14419 pmid: 28098947 |
| [21] |
Goslin K, Zheng B B, Serrano-Mislata A, Rae L, Ryan P T, Kwaśniewska K, Thomson B, Ó’Maoiléidigh D S, Madueño F, Wellmer F, Graciet E. Transcription factor interplay between LEAFY and APETALA1/CAULIFLOWER during floral initiation. Plant Physiol, 2017, 174: 1097-1109.
doi: 10.1104/pp.17.00098 pmid: 28385730 |
| [22] |
Han Y Y, Zhang C, Yang H B, Jiao Y L. Cytokinin pathway mediates APETALA1 function in the establishment of determinate floral meristems in Arabidopsis. Proc Natl Acad Sci USA, 2014, 111: 6840-6845.
doi: 10.1073/pnas.1318532111 |
| [23] |
Kim W C, Ko J H, Kim J Y, Kim J, Bae H J, Han K H. MYB 46directly regulates the gene expression of secondary wall- associated cellulose synthases in Arabidopsis. Plant J, 2013, 73: 26-36.
doi: 10.1111/j.1365-313x.2012.05124.x |
| [24] |
Kim W C, Kim J Y, Ko J H, Kim J, Han K H. Transcription factor MYB46 is an obligate component of the transcriptional regulatory complex for functional expression of secondary wall- associated cellulose synthases in Arabidopsis thaliana . J Plant Physiol, 2013, 170: 1374-1378.
doi: 10.1016/j.jplph.2013.04.012 |
| [25] |
Zhong R, Ye Z H. MYB46 and MYB83 bind to the SMRE sites and directly activate a suite of transcription factors and secondary wall biosynthetic genes. Plant Cell Physiol, 2012, 53: 368-380.
doi: 10.1093/pcp/pcr185 |
| [26] |
Wang X C, Wu J, Guan M L, Zhao C H, Geng P, Zhao Q. Arabidopsis MYB4 plays dual roles in flavonoid biosynthesis. Plant J, 2020, 101: 637-652.
doi: 10.1111/tpj.14570 |
| [27] |
Lazakis C M, Coneva V, Colasanti J. ZCN8 encodes a potential orthologue of Arabidopsis FT florigen that integrates both endogenous and photoperiod flowering signals in maize. J Exp Bot, 2011, 62: 4833-4842.
doi: 10.1093/jxb/err129 pmid: 21730358 |
| [28] |
Coneva V, Guevara D, Rothstein S J, Colasanti J. Transcript and metabolite signature of maize source leaves suggests a link between transitory starch to sucrose balance and the autonomous floral transition. J Exp Bot, 2012, 63: 5079-5092.
doi: 10.1093/jxb/ers158 |
| [29] |
Mara C D, Huang T, Irish V F. The Arabidopsis floral homeotic proteins APETALA3 and PISTILLATA negatively regulate the BANQUO genes implicated in light signaling. Plant Cell, 2010, 22: 690-702.
doi: 10.1105/tpc.109.065946 |
| [30] |
Kim K C, Lai Z, Fan B, Chen Z. Arabidopsis WRKY38 and WRKY62 transcription factors interact with histone deacetylase 19 in basal defense. Plant Cell, 2008, 20: 2357-2371.
doi: 10.1105/tpc.107.055566 |
| [31] |
Mao P, Duan M R, Wei C H, Li Y. WRKY62 transcription factor acts downstream of cytosolic NPR1 and negatively regulates jasmonate-responsive gene expression. Plant Cell Physiol, 2007, 48: 833-842.
doi: 10.1093/pcp/pcm058 |
| [32] |
Grunewald W, De Smet I, Lewis D R, Löfke C, Jansen L, Goeminne G, Vanden Bossche R, Karimi M, De Rybel B, Vanholme B, Teichmann T, Boerjan W, Van Montagu M C, Gheysen G, Muday G K, Friml J, Beeckman T. Transcription factor WRKY23 assists auxin distribution patterns during Arabidopsis root development through local control on flavonol biosynthesis. Proc Natl Acad Sci USA, 2012, 109: 1554-1559.
doi: 10.1073/pnas.1121134109 |
| [33] |
Prát T, Hajný J, Grunewald W, Vasileva M, Molnár G, Tejos R, Schmid M, Sauer M, Friml J. WRKY23 is a component of the transcriptional network mediating auxin feedback on PIN polarity. PLoS Genet, 2018, 14: e1007177.
doi: 10.1371/journal.pgen.1007177 |
| [34] |
Guo X Y, Wang Y, Zhao P X, Xu P, Yu G H, Zhang L Y, Xiong Y, Xiang C B. AtEDT1/HDG11 regulates stomatal density and water-use efficiency via ERECTA and E2Fa. New Phytol, 2019, 223: 1478-1488.
doi: 10.1111/nph.15861 |
| [35] |
Cai X T, Xu P, Wang Y, Xiang C B. Activated expression of AtEDT1/HDG11 promotes lateral root formation in Arabidopsis mutant edt1 by upregulating jasmonate biosynthesis. J Integr Plant Biol, 2015, 57: 1017-1730.
doi: 10.1111/jipb.12347 |
| [36] |
Yu L H, Wu S J, Peng Y S, Liu R N, Chen X, Zhao P, Xu P, Zhu J B, Jiao G L, Pei Y, Xiang C B. Arabidopsis EDT1/HDG11 improves drought and salt tolerance in cotton and poplar and increases cotton yield in the field. Plant Biotechnol J, 2016, 14: 72-84.
doi: 10.1111/pbi.12358 |
| [37] |
Vanderauwera S, Vandenbroucke K, Inzé A, van de Cotte B, Mühlenbock P, De Rycke R, Naouar N, Van Gaever T, Van Montagu M C, Van Breusegem F. AtWRKY 15perturbation abolishes the mitochondrial stress response that steers osmotic stress tolerance in Arabidopsis. Proc Natl Acad Sci USA, 2012, 109: 20113-20118.
doi: 10.1073/pnas.1217516109 |
| [38] |
Ge S T, Han X F, Xu X W, Shao Y M, Zhu Q K, Liu Y D, Du J, Xu J, Zhang S Q. WRKY15 suppresses tracheary element differentiation upstream of VND7 during xylem formation. Plant Cell, 2020, 32: 2307-2324.
doi: 10.1105/tpc.19.00689 |
| [39] |
Gu Z Y, Men S Q, Zhu J, Hao Q, Tong N N, Liu Z A, Zhang H C, Shu Q Y, Wang L S. Chalcone synthase is ubiquitinated and degraded via interactions with a RING-H2 protein in petals of Paeonia‘he xie’. J Exp Bot, 2019, 70: 4749-4762.
doi: 10.1093/jxb/erz245 |
| [40] | 张丽玲, 郄倩茹, 罗韶凡, 牛文康, 朱喆标, 高雨柔, 李旭凯, 韩渊怀. 谷子12种黄酮类代谢物合成通路分析. 山西农业大学学报(自然科学版), 2020, 40(4): 10-18. |
| Zhang L L, Qie Q R, Luo S F, Niu W K, Zhu Z B, Gao Y R, Li X K, Han Y H. Analysis of synthesis pathway of twelve flavonoid metabolites in foxtail millet. J Shanxi Agric Univ (Nat Sci Edn), 2020, 40(4): 10-18. (in Chinese with English abstract) | |
| [41] |
Gonzalez A, Zhao M Z, Leavitt J M, Lloyd A M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J, 2008, 53: 814-827.
doi: 10.1111/j.1365-313X.2007.03373.x |
| [42] |
Mohamed H I, Latif H H. Improvement of drought tolerance of soybean plants by using methyl jasmonate. Physiol Mol Biol Plants, 2017, 23: 545-556.
doi: 10.1007/s12298-017-0451-x |
| [1] | 王蓉, 陈小红, 王倩, 刘少雄, 陆平, 刁现民, 刘敏轩, 王瑞云. 中国谷子名米品种遗传多样性与亲缘关系研究[J]. 作物学报, 2022, 48(8): 1914-1925. |
| [2] | 李佩婷, 赵振丽, 黄潮华, 黄国强, 徐良年, 邓祖湖, 张玉, 赵新旺. 基于转录组及WGCNA的甘蔗干旱响应调控网络分析[J]. 作物学报, 2022, 48(7): 1583-1600. |
| [3] | 柯丹霞, 霍娅娅, 刘怡, 李锦颖, 刘晓雪. 大豆TGA转录因子基因GmTGA26在盐胁迫中的功能分析[J]. 作物学报, 2022, 48(7): 1697-1708. |
| [4] | 郭楠楠, 刘天策, 史硕, 胡心亭, 牛亚丹, 李亮. 长链非编码RNA (LncRNA)在印度梨形孢促进大麦根部生长发育中的调控作用[J]. 作物学报, 2022, 48(7): 1625-1634. |
| [5] | 朱峥, 王田幸子, 陈悦, 刘玉晴, 燕高伟, 徐珊, 马金姣, 窦世娟, 李莉云, 刘国振. 水稻转录因子WRKY68在Xa21介导的抗白叶枯病反应中发挥正调控作用[J]. 作物学报, 2022, 48(5): 1129-1140. |
| [6] | 黄伟, 高国应, 吴金锋, 刘丽莉, 张大为, 周定港, 成洪涛, 张凯旋, 周美亮, 李莓, 严明理. 芥菜型油菜BjA09.TT8和BjB08.TT8基因调节类黄酮的合成[J]. 作物学报, 2022, 48(5): 1169-1180. |
| [7] | 陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展[J]. 作物学报, 2022, 48(4): 781-790. |
| [8] | 晋敏姗, 曲瑞芳, 李红英, 韩彦卿, 马芳芳, 韩渊怀, 邢国芳. 谷子糖转运蛋白基因SiSTPs的鉴定及其参与谷子抗逆胁迫响应的研究[J]. 作物学报, 2022, 48(4): 825-839. |
| [9] | 杜晓芬, 王智兰, 韩康妮, 连世超, 李禹欣, 张林义, 王军. 谷子叶绿体基因RNA编辑位点的鉴定与分析[J]. 作物学报, 2022, 48(4): 873-885. |
| [10] | 赵美丞, 刁现民. 谷子近缘野生种的亲缘关系及其利用研究[J]. 作物学报, 2022, 48(2): 267-279. |
| [11] | 尹明, 杨大为, 唐慧娟, 潘根, 李德芳, 赵立宁, 黄思齐. 大麻GRAS转录因子家族的全基因组鉴定及镉胁迫下表达分析[J]. 作物学报, 2021, 47(6): 1054-1069. |
| [12] | 葛敏, 王元琮, 宁丽华, 胡梦梅, 石习, 赵涵. 氮响应转录因子ZmNLP5影响玉米根系生长的功能研究[J]. 作物学报, 2021, 47(5): 807-813. |
| [13] | 马贵芳, 满夏夏, 张益娟, 高豪, 孙朝霞, 李红英, 韩渊怀, 侯思宇. 谷子穗发育期转录组与叶酸代谢谱联合分析[J]. 作物学报, 2021, 47(5): 837-846. |
| [14] | 贾小平, 李剑峰, 张博, 全建章, 王永芳, 赵渊, 张小梅, 王振山, 桑璐曼, 董志平. 谷子SiPRR37基因对光温、非生物胁迫的响应特点及其有利等位变异鉴定[J]. 作物学报, 2021, 47(4): 638-649. |
| [15] | 邱红梅, 陈亮, 侯云龙, 王新风, 陈健, 马晓萍, 崔正果, 张玲, 胡金海, 王跃强, 邱丽娟. 大豆种子颜色遗传调控机制研究进展[J]. 作物学报, 2021, 47(12): 2299-2313. |
|