欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (8): 1948-1956.doi: 10.3724/SP.J.1006.2022.12049
王穆穆1( ), 何艳芳2, 郑永胜1, 王晖1, 王丽媛1, 王东建1, 张晗1, 李汝玉1,*()
), 何艳芳2, 郑永胜1, 王晖1, 王丽媛1, 王东建1, 张晗1, 李汝玉1,*()
WANG Mu-Mu1(), HE Yan-Fan2, ZHENG Yong-Sheng1, WANG Hui1, WANG Li-Yuan1, WANG Dong-Jian1, ZHANG Han1, LI Ru-Yu1,*()
摘要:
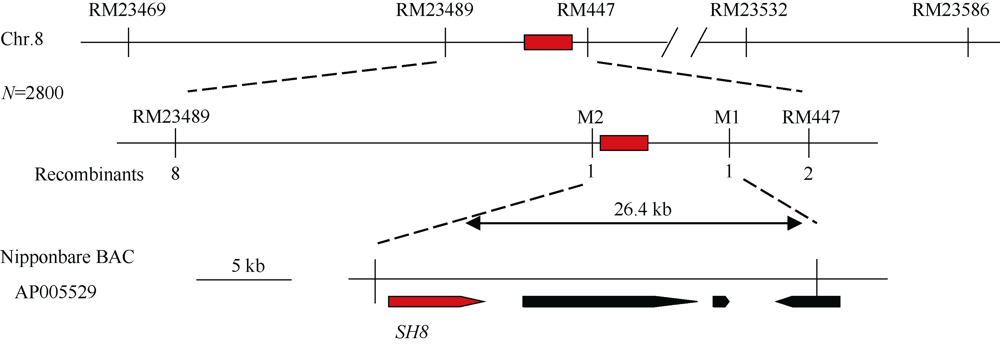
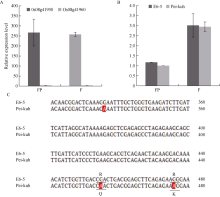
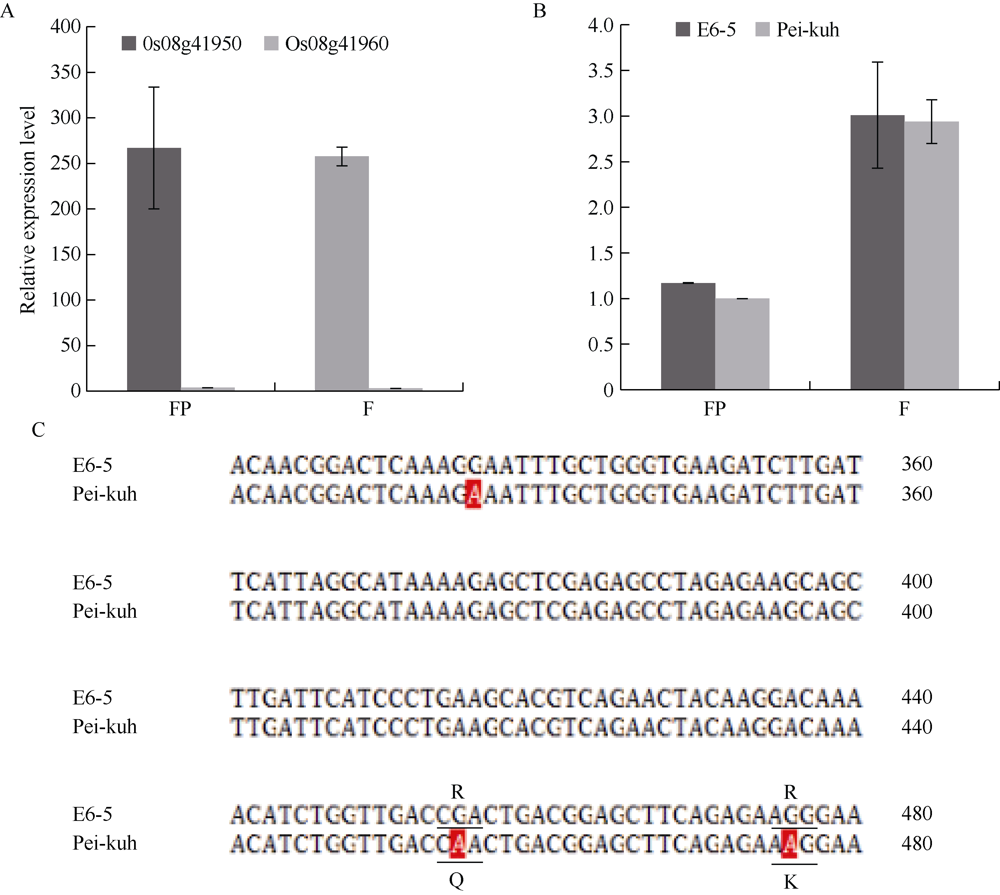
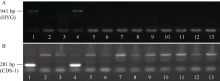
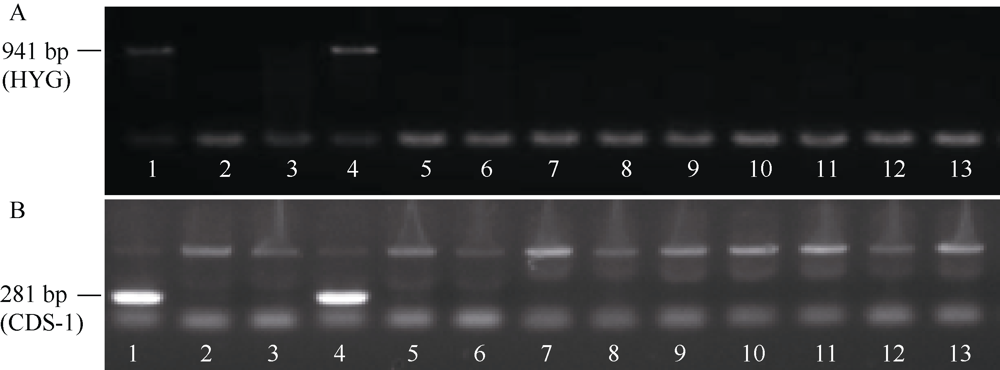
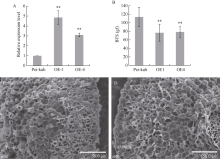
落粒性是水稻的重要性状之一, 是野生稻种子传播和后代繁衍的重要保障, 但是容易落粒在栽培稻的生产中会造成产量损失。目前, 在水稻中已经克隆了一些落粒相关基因, 但是这些基因并不能解释水稻落粒的全部表型变异。本研究从白谷(Pei-kuh)和普通野生稻(W1944)构建的重组自交系中选取落粒性强且不含有SH4基因的材料E6-5, 与Pei-kuh进行杂交构建分离群体, 将以前检测到位于8号染色体的落粒QTL精细定位于26.4 kb的区间内, 并把该位点的落粒基因命名为SHATTERING8 (SH8)。该区段共有4个编码基因, Pei-kuh和E6-5的测序比对发现LOC_Os08g41950基因在编码区有3个碱基的差异, 同时实时定量PCR分析表明该基因在离层部位表达, 因此我们把LOC_Os08g41950作为SH8的目标候选基因。为了验证LOC_Os08g41950是否参与水稻落粒的调控, 我们利用来自E6-5的LOC_Os08g41950编码序列构建了过表达载体并遗传转化Pei-huh。落粒表型鉴定结果表明, 与Pei-huh相比, 转基因植株的落粒性明显增强, 不仅种子脱落所需的拉力值显著降低, 而且还具有更光滑的离层断裂面。本研究有助于促进水稻驯化和水稻落粒调控机制的研究。
| [1] |
Doebley J F, Gaut B S, Smith B D. The molecular genetics of crop domestication. Cell, 2006, 127: 1309-1321.
pmid: 17190597 |
| [2] |
Fuller D Q, Qin L, Zheng Y, Zhao Z, Chen X, Hosoya L A, Sun G P. The domestication process and domestication rate in rice: spikelet bases from the Lower Yangtze. Science, 2009, 323: 1607-1610.
doi: 10.1126/science.1166605 |
| [3] |
Taylor J E, Whitelaw C A. Signals in abscission. New Phytol, 2001, 151: 323-340.
doi: 10.1046/j.0028-646x.2001.00194.x |
| [4] |
Taghizadeh M S, Simon C, Nicolas M E, Cousens R D. Water deficit changes the anatomy of the fruit abscission zone in Raphanus raphanistrum (Brassicaceae). Aust J Bot, 2009, 57: 708-714.
doi: 10.1071/BT09165 |
| [5] |
Zhou Y, Lu D, Li C, Luo J, Zhu B F, Zhu J, Shang-Guan Y, Wang Z, Sang T, Zhou B, Han B. Genetic control of seed shattering in rice by the APETALA2 transcription factor shattering abortion1. Plant Cell, 2012, 24: 1034-1048.
doi: 10.1105/tpc.111.094383 |
| [6] |
Xiong Z L, Liu D K, Dai K X, Xu G C, Zhang Q. Identification of genetic factors controlling domestication-related traits of rice using an F2 population of a cross between Oryza sativa and O. rufipogon. Theor Appl Genet, 1999, 98: 243-251.
doi: 10.1007/s001220051064 |
| [7] |
Cai H W, Morishima H. Genomic regions affecting seed shattering and seed dormancy in rice. Theor Appl Genet, 2000, 100: 840-846.
doi: 10.1007/s001220051360 |
| [8] |
Thomson M J, Tai T H, McClung A M, Lai X H, Hinga M E, Lobos K B, Xu Y, Martinez C P, McCouch S R. Mapping quantitative trait loci for yield, yield components and morphological traits in an advanced backcross population between Oryza rufipogon and the Oryza sativa cultivar Jefferson. Theor Appl Genet, 2003, 107: 479-493.
pmid: 12736777 |
| [9] |
Gu X Y, Kianian S F, Hareland G A, Hoffer B L, Foley M E. Genetic analysis of adaptive syndromes interrelated with seed dormancy in weedy rice (Oryza sativa). Theor Appl Genet, 2005, 110: 1108-1118.
doi: 10.1007/s00122-005-1939-2 |
| [10] | Li C, Zhou A, Sang T. Genetic analysis of rice domestication syndrome with the wild annual species, Oryza nivara. New Phytol, 2006, 170: 185-193. |
| [11] |
Konishi S, Izawa T, Lin S Y, Ebana K, Fukuta Y, Sasaki T, Yano M. An SNP caused loss of seed shattering during rice domestication. Science, 2006, 312: 1392-1396.
pmid: 16614172 |
| [12] |
Li C, Zhou A, Sang T. Rice domestication by reducing shattering. Science, 2006, 311: 1936-1939.
doi: 10.1126/science.1123604 |
| [13] |
Lin Z, Griffith M E, Li X, Zhu Z, Tan L, Fu Y, Zhang W, Wang X, Xie D, Sun C. Origin of seed shattering in rice (Oryza sativa L.). Planta, 2007, 226: 11-20.
doi: 10.1007/s00425-006-0460-4 |
| [14] |
Yoon J, Cho L H, Kim S L, Choi H, Koh H J, An G. The BEL1-type homeobox gene SH5 induces seed shattering by enhancing abscission-zone development and inhibiting lignin biosynthesis. Plant J, 2014, 79: 717-728.
doi: 10.1111/tpj.12581 |
| [15] |
Ji H, Kim S R, Kim Y H, Kim H, Eun M Y, Jin I D, Cha Y S, Yun D W, Ahn B O, Lee M C, Lee G S, Yoon U H, Lee J S, Lee Y H, Suh S C, Jiang W, Yang J I, Jin P, McCouch S R, An G, Koh H J. Inactivation of the CTD phosphatase-like gene OsCPL1 enhances the development of the abscission layer and seed shattering in rice. Plant J, 2010, 61: 96-106.
doi: 10.1111/j.1365-313X.2009.04039.x |
| [16] |
Yoon J, Cho L H, Antt H W, Koh H J, An G. KNOX protein OSH15 induces grain shattering by repressing lignin biosynthesis genes. Plant Physiol, 2017, 174: 312-325.
doi: 10.1104/pp.17.00298 |
| [17] |
Wu W, Liu X, Wang M, Meyer R S, Luo X, Ndjiondjop M N, Tan L, Zhang J, Wu J, Cai H, Sun C, Wang X, Wing R A, Zhu Z. A single-nucleotide polymorphism causes smaller grain size and loss of seed shattering during African rice domestication. Nat Plants, 2017, 3: 17064.
doi: 10.1038/nplants.2017.64 |
| [18] |
Lyu S, Wu W, Wang M, Meyer R S, Ndjiondjop M N, Tan L, Zhou H, Zhang J, Fu Y, Cai H, Sun C, Wing R A, Zhu Z. Genetic control of seed shattering during African rice domestication. Nat Plants, 2018, 4: 331-337.
doi: 10.1038/s41477-018-0164-3 |
| [19] |
Jiang L, Ma X, Zhao S, Tang Y, Liu F, Gu P, Fu Y, Zhu Z, Cai H, Sun C, Tan L. The APETALA2-Like transcription factor SUPERNUMERARY BRACT controls rice seed shattering and seed size. Plant Cell, 2019, 31: 17-36.
doi: 10.1105/tpc.18.00304 |
| [20] |
Ji H S, Chu S H, Jiang W, Cho Y I, Hahn J H, Eun M Y, McCouch S R, Koh H J. Characterization and mapping of a shattering mutant in rice that corresponds to a block of domestication genes. Genetics, 2006, 173: 995-1005.
doi: 10.1534/genetics.105.054031 |
| [21] |
Liljegren S J, Ditta G S, Eshed Y, Savidge B, Bowman J L, Yanofsky M F. SHATTERPROOF MADS-box genes control seed dispersal in Arabidopsis. Nature, 2000, 404: 766-770.
doi: 10.1038/35008089 |
| [22] |
Fernandez D E, Heck G R, Perry S E, Patterson S E, Bleecker A B, Fang S C. The embryo MADS domain factor AGL15 acts postembryonically. Inhibition of perianth senescence and abscission via constitutive expression. Plant Cell, 2000, 12: 183-198.
pmid: 10662856 |
| [23] |
Mao L, Begum D, Chuang H W, Budiman M A, Szymkowiak E J, Irish E E, Wing R A. JOINTLESS is a MADS-box gene controlling tomato flower abscissionzone development. Nature, 2000, 406: 910-913.
doi: 10.1038/35022611 |
| [24] | Nakano T, Kimbara J, Fujisawa M, Kitagawa M, Ihashi N, Maeda H, Kasumi T, Ito Y. MACROCALYX and JOINTLESS interact in the transcriptional regulation of tomato fruit abscission zone development. Plant Physiol, 2012, 158: 439-450. |
| [25] |
Liu D, Wang D, Qin Z, Zhang D, Yin L, Wu L, Colasanti J, Li A, Mao L. The SEPALLATA MADS-box protein SLMBP21 forms protein complexes with JOINTLESS and MACROCALYX as a transcription activator for development of the tomato flower abscission zone. Plant J, 2014, 77: 284-296.
doi: 10.1111/tpj.12387 |
| [26] |
Onishi K, Takagi K, Kontani M, Tanaka T, Sano Y. Different patterns of genealogical relationships found in the two major QTLs causing reduction of seed shattering during rice domestication. Genome, 2007, 50: 757-766.
doi: 10.1139/G07-051 |
| [27] |
Favaro R, Pinyopich A, Battaglia R, Kooiker M, Borghi L, Ditta G, Yanofsky M F, Kater M M, Colombo L. MADS-Box protein complexes control carpel and ovule development in Arabidopsis. Plant Cell, 2003, 15: 2603-2611.
doi: 10.1105/tpc.015123 |
| [28] |
Kater M M, Dreni L, Colombo L. Functional conservation of MADS-box factors controlling floral organ identity in rice and Arabidopsis. J Exp Bot, 2006, 57: 3433-3444.
doi: 10.1093/jxb/erl097 |
| [1] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [2] | 周慧文, 丘立杭, 黄杏, 李强, 陈荣发, 范业赓, 罗含敏, 闫海锋, 翁梦苓, 周忠凤, 吴建明. 甘蔗赤霉素氧化酶基因ScGA20ox1的克隆及功能分析[J]. 作物学报, 2022, 48(4): 1017-1026. |
| [3] | 谢琴琴, 左同鸿, 胡燈科, 刘倩莹, 张以忠, 张贺翠, 曾文艺, 袁崇墨, 朱利泉. 甘蓝自交不亲和相关基因BoPUB9的克隆及表达分析[J]. 作物学报, 2022, 48(1): 108-120. |
| [4] | 罗兰, 雷丽霞, 刘进, 张瑞华, 金桂秀, 崔迪, 黎毛毛, 马小定, 赵正武, 韩龙植. 利用东乡普通野生稻染色体片段置换系定位产量相关性状QTL[J]. 作物学报, 2021, 47(7): 1391-1401. |
| [5] | 唐锐敏, 贾小云, 朱文娇, 印敬明, 杨清. 马铃薯热激转录因子HsfA3基因的克隆及其耐热性功能分析[J]. 作物学报, 2021, 47(4): 672-683. |
| [6] | 岳洁茹, 白建芳, 张风廷, 郭丽萍, 苑少华, 李艳梅, 张胜全, 赵昌平, 张立平. 杂交小麦抗坏血酸过氧化物酶基因克隆及其在种子老化中的潜在功能分析[J]. 作物学报, 2021, 47(3): 405-415. |
| [7] | 杨琴莉, 杨多凤, 丁林云, 赵汀, 张军, 梅欢, 黄楚珺, 高阳, 叶莉, 高梦涛, 严孙艺, 张天真, 胡艳. 棉花花器官突变体的鉴定及候选基因的克隆[J]. 作物学报, 2021, 47(10): 1854-1862. |
| [8] | 何潇, 刘兴, 辛正琦, 谢海艳, 辛余凤, 吴能表. 半夏PtPAL基因的克隆、表达与酶动力学分析[J]. 作物学报, 2021, 47(10): 1941-1952. |
| [9] | 左同鸿, 张贺翠, 刘倩莹, 廉小平, 谢琴琴, 胡燈科, 张以忠, 王玉奎, 白晓璟, 朱利泉. 甘蓝自交不亲和性相关基因BoGSTL21的克隆与表达分析[J]. 作物学报, 2020, 46(12): 1850-1861. |
| [10] | 冯韬,官春云. 甘蓝型油菜光敏色素互作因子4 (BnaPIF4)基因克隆和功能分析[J]. 作物学报, 2019, 45(2): 204-213. |
| [11] | 薛晓梦,李建国,白冬梅,晏立英,万丽云,康彦平,淮东欣,雷永,廖伯寿. 花生FAD2基因家族表达分析及其对低温胁迫的响应[J]. 作物学报, 2019, 45(10): 1586-1594. |
| [12] | 谈欢,刘玉汇,李丽霞,王丽,李元铭,张俊莲. 马铃薯块茎花色素苷合成相关R2R3 MYB蛋白基因的克隆和功能 分析[J]. 作物学报, 2018, 44(7): 1021-1031. |
| [13] | 冯韬,官春云. 甘蓝型油菜芸薹素唑耐受因子(BnaBZR1/BnaBES1)全长CDS克隆与生物信息学分析[J]. 作物学报, 2018, 44(12): 1793-1801. |
| [14] | 马晨雨,詹为民,李文亮,张梦迪,席章营. 玉米ZmNAOD基因的克隆与功能分析[J]. 作物学报, 2018, 44(10): 1433-1441. |
| [15] | 刘朝显, 王久光, 梅秀鹏, 余婷婷, 王国强, 周练, 蔡一林. 玉米胚乳母本印记基因ZmVIL1的克隆及印记特性分析[J]. 作物学报, 2018, 44(03): 376-384. |
|