欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (4): 672-683.doi: 10.3724/SP.J.1006.2021.04114
唐锐敏1,2( ), 贾小云1, 朱文娇2, 印敬明2, 杨清2,*()
), 贾小云1, 朱文娇2, 印敬明2, 杨清2,*()
TANG Rui-Min1,2(), JIA Xiao-Yun1, ZHU Wen-Jiao2, YIN Jing-Ming2, YANG Qing2,*()
摘要:
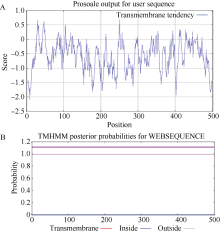
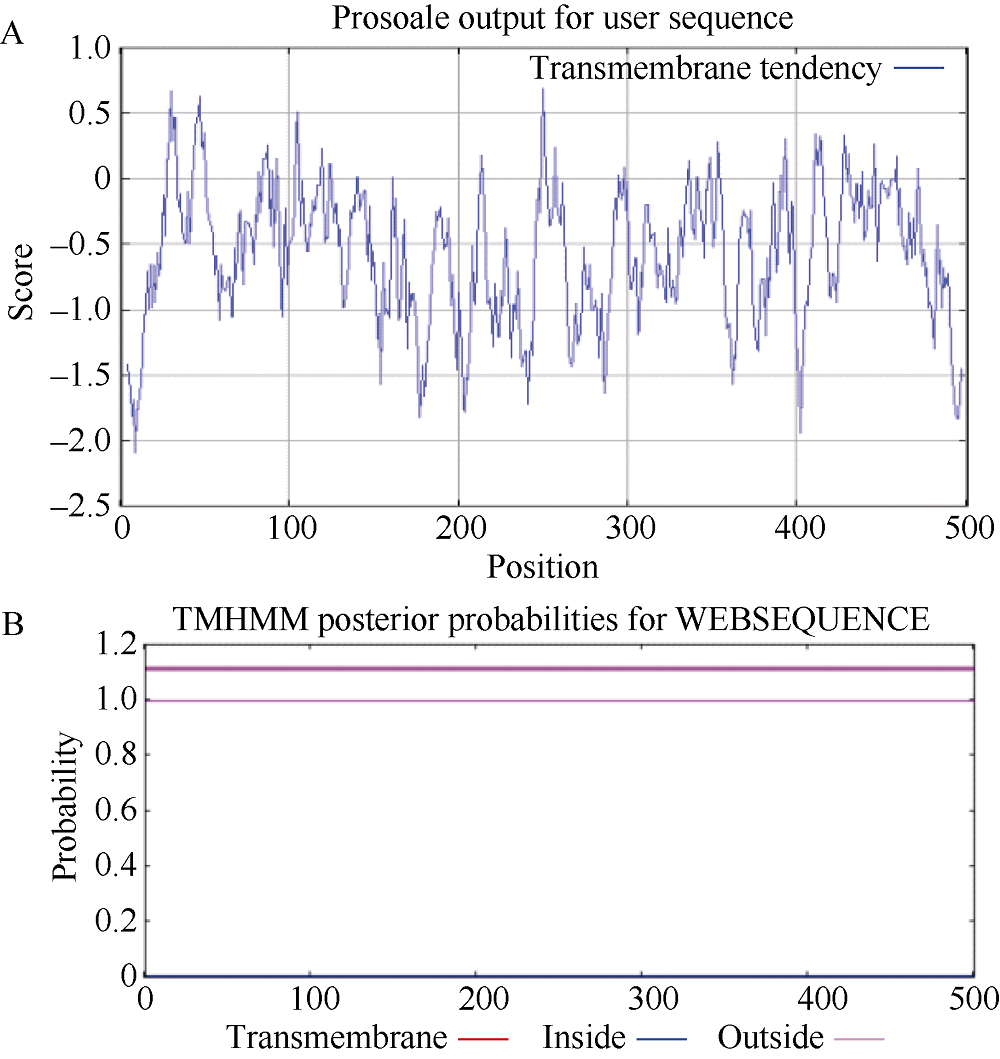
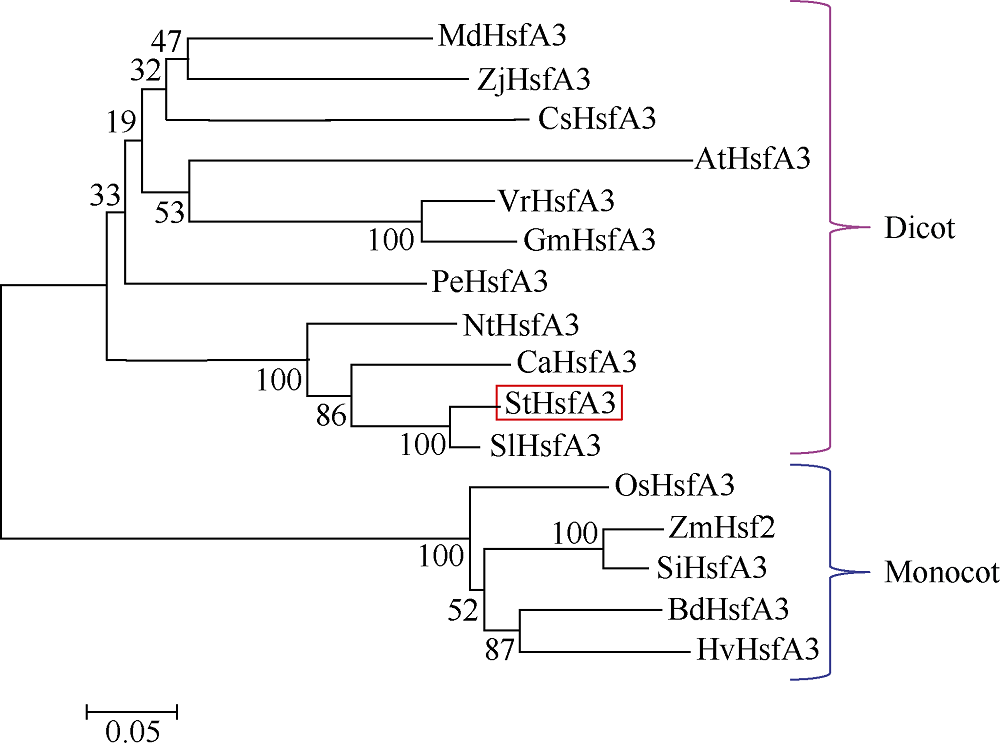
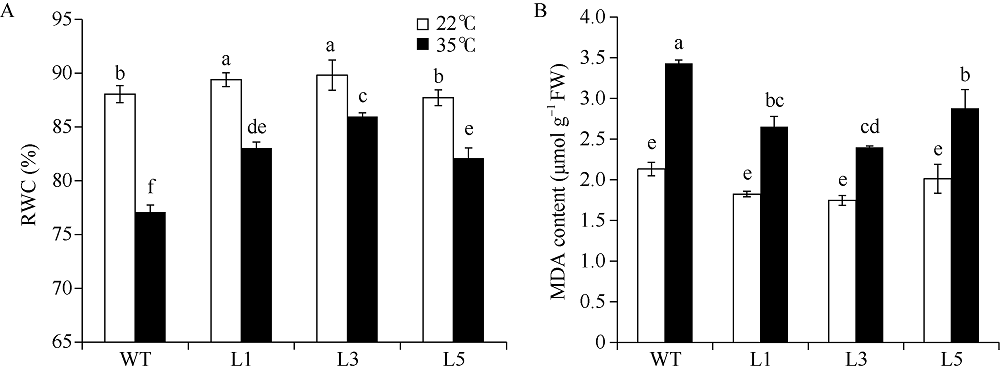
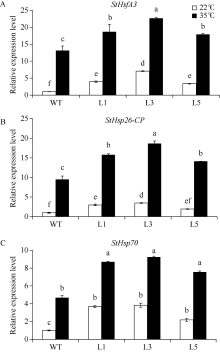
马铃薯在田间生长时常会受到各种不利环境的影响。夏季高温常导致马铃薯块茎产量和质量的下降。因此, 阐明马铃薯对热胁迫的响应机制, 发掘耐热相关基因, 对马铃薯耐热性的提高意义重大。热激转录因子(heat shock transcription factor A3, HsfA3)在植物机体内的活动影响到大量功能基因的表达, 在植物响应热胁迫的过程中发挥重要的作用。为了研究马铃薯中HsfA3的结构和功能, 本研究通过RT-PCR从马铃薯品种Désirée中克隆到长度为1506 bp的StHsfA3基因, 编码501个氨基酸。StHsfA3的相对分子量为55.23 kD, 理论等电点为4.9, 属于亲水性蛋白。构建StHsfA3-pBA002过表达载体, 并转化马铃薯植株, 共鉴定得到5个独立的StHsfA3过表达转基因马铃薯株系。通过对转基因植株和非转基因植株叶片中的相对含水量(relative water content, RWC)以及丙二醛(malondialdehyde, MDA)含量的测定发现, 在高温胁迫下, 转基因植株叶片中的RWC显著高于非转基因植株, MDA含量显著低于非转基因植株, 说明StHsfA3在耐热过程中起到正调控作用。对StHsfA3、StHsp26-CP和StHsp70在不同马铃薯植株中的表达分析显示, StHsfA3过量表达诱导了StHsp26-CP和StHsp70的表达, 预示着StHsfA3可能协同StHsp26-CP和StHsp70来增强过表达转基因株系的耐热性。
| [1] | Zhu J K. Abiotic stress signaling and responses in plants. Cell, 2016,167:313-324. |
| [2] | Herman D J, Knowles L O, Knowles N R. Heat stress affects carbohydrate metabolism during cold-induced sweetening of potato ( Solanum tuberosum L.). Planta, 2017,245:563-582. |
| [3] | Lafta A, Lorenzen J. Effect of high temperature on plant growth and carbohydrate metabolism in potato. Plant Physiol, 1995,109:637-643. |
| [4] |
Kotak S, Larkindale J, Lee U, von Koskull-Döring P, Vierling E, Scharf K D. Complexity of the heat stress response in plants. Curr Opin Plant Biol, 2007,10:310-316.
doi: 10.1016/j.pbi.2007.04.011 pmid: 17482504 |
| [5] | Scharf K D, Berberich T, Ebersberger I, Nover L. The plant heat stress transcription factor (Hsf) family: structure, function and evolution. Biochim Biophys Acta, 2012,1819:104-119. |
| [6] | Lyck R, Harmening U, Höhfeld I, Treuter E, Scharf K D, Nover L. Intracellular distribution and identification of the nuclear localization signals of two plant heat-stress transcription factors. Planta, 1997,202:117-125. |
| [7] | Miller G, Mittler R. Could heat shock transcription factors function as hydrogen peroxide sensors in plants? Ann Bot, 2006,98:279-288. |
| [8] | Nover L, Bharti K, Döring P, Mishra S K, Ganguli A, Scharf K D. Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need? Cell Stress Chaperones, 2001,6:177-189. |
| [9] | Baniwal S K, Bharti K, Chan K Y, Fauth M, Ganguli A, Kotak S, Mishra S K, Nover L, Port M, Scharf K D, Tripp J, Weber C, Zielinski D, von Koskull-Döring P. Heat stress response in plants: a complex game with chaperones and more than twenty heat stress transcription factors. J Biosci, 2004,29:471-487. |
| [10] |
Guo J, Wu J, Ji Q, Wang C, Luo L, Yuan Y, Wang Y, Wang J. Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis. J Genet Genomics, 2008,35:105-118.
pmid: 18407058 |
| [11] | Wang F, Dong Q, Jiang H, Zhu S, Chen B, Xiang Y. Genome-wide analysis of the heat shock transcription factors in Populus trichocarpa and Medicago truncatula. Mol Biol Rep, 2012,39:1877-1886. |
| [12] | Bharti K, Koskull-Döring P V, Bharti S, Kumar P, Tintschl-Körbitzer A, Treuter E, Nover L. Tomato heat stress transcription factor HsfB1 represents a novel type of general transcription coactivator with a histone-like motif interacting with the plant CREB binding protein ortholog HAC1. Plant Cell, 2004,16:1521-1535. |
| [13] | Swindell W R, Huebner M, Weber A P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Genomics, 2007,8:125. |
| [14] | Zhang J, Liu B, Li J, Zhang L, Wang Y, Zheng H, Lu M, Chen J. Hsf and Hsp gene families in Populus: genome-wide identification, organization and correlated expression during development and in stress responses. BMC Genomics, 2015,16:181. |
| [15] |
Tang R M, Zhu W J, Song X Y, Lin X Z, Cai J H, Wang M, Yang Q. Genome-wide identification and function analyses of heat shock transcription factors in potato. Front Plant Sci, 2016,7:490.
pmid: 27148315 |
| [16] | Bharti K, Schmidt E, Lyck R, Heerklotz D, Bublak D, Scharf K D. Isolation and characterization of HsfA3, a new heat stress transcription factor of Lycopersicon peruvianum. Plant J, 2000,22:355-365. |
| [17] | Yoshida T, Sakuma Y, Todaka D, Maruyama K, Qin F, Mizoi J, Kidokoro S, Fujita Y, Shinozaki K, Yamaguchi-Shinozaki K. Functional analysis of an Arabidopsis heat-shock transcription factor HsfA3 in the transcriptional cascade downstream of the DREB2A stress-regulatory system. Biochem Biophys Res Commun, 2008,368:515-521. |
| [18] | Link V, Sinha A K, Vashista P, Hofmann M G, Proels R K, Ehness R, Roitsch T. A heat-activated MAP kinase in tomato: a possible regulator of the heat stress response. FEBS Lett, 2002,531:179-183. |
| [19] | Zhu M D, Zhang M, Gao D J, Zhou K, Tang S J, Zhou B, Lyu Y M. Rice OsHSFA3 gene improves drought tolerance by modulating polyamine biosynthesis depending on abscisic acid and ROS levels. Int J Mol Sci, 2020,21:E1857. |
| [20] |
Wu Z, Liang J, Wang C, Zhao X, Zhong X, Cao X, Li G, He J, Yi M. Overexpression of lily HsfA3s in Arabidopsis confers increased thermotolerance and salt sensitivity via alterations in proline catabolism. J Exp Bot, 2018,69:2005-2021.
pmid: 29394377 |
| [21] |
Li W, Wang B, Wang M, Chen M, Yin J M, Kaleri G M, Zhang R J, Zuo T N, You X, Yang Q. Cloning and characterization of a potato StAN11 gene involved in anthocyanin biosynthesis regulation. J Integr Plant Biol, 2014,56:364-372.
doi: 10.1111/jipb.12136 pmid: 24304603 |
| [22] | Jin Q, Zhu K, Cui W, Xie Y, Han B, Shen W. Hydrogen gas acts as a novel bioactive molecule in enhancing plant tolerance to paraquat-induced oxidative stress via the modulation of hemeoxygenase-1 signalling system. Plant Cell Environ, 2013,36:956-969. |
| [23] | Rykaczewska K. The impact of high temperature during growing season on potato cultivars with different response to environmental stresses. Am J Plant Sci, 2013,4:2386-2393. |
| [24] | Tang R M, Niu S Y, Zhang G D, Chen G S, Haroon M, Yang Q, Rajora O P, Li X Q. Physiological and growth responses of potato cultivars to heat stress. Botany, 2018,96:897-912. |
| [25] | Ikeda M, Ohme-Takagi M. A novel group of transcriptional repressors in Arabidopsis. Plant Cell Physiol, 2009,50:970-975. |
| [26] |
Li P S, Yu T F, He G H, Chen M, Zhou Y B, Chai S C, Xu Z S, Ma Y Z. Genome-wide analysis of the Hsf family in soybean and functional identification of GmHsf-34 involvement in drought and heat stresses. BMC Genomics, 2014,15:1009.
pmid: 25416131 |
| [27] |
Emani C, Garcia J M, Lopata-Finch E, Pozo M J, Uribe P, Kim D J, Sunilkumar G, Cook D R, Kenerley C M, Rathore K S. Enhanced fungal resistance in transgenic cotton expressing an endochitinase gene from Trichoderma virens. Plant Biotechnol J, 2003,1:321-336.
doi: 10.1046/j.1467-7652.2003.00029.x pmid: 17166131 |
| [28] | 贾小霞, 齐恩芳, 刘石, 文国宏, 马胜, 李建武, 黄伟. AtDREB1A基因过量表达对马铃薯生长及抗非生物胁迫基因表达的影响. 作物学报, 2019,45:1166-1175. |
| Jia X X, Qi E F, Liu S, Wen G H, Ma S, Li J W, Huang W. Effects of over-expression of AtDREB1A gene on potato growth and abiotic stress resistance gene expression. Acta Agron Sin, 2019,45:1166-1175. (in Chinese with English abstract). | |
| [29] | Gawel S, Wardas M, Niedworok E, Wardas P. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad Lek, 2004,57:453-455. |
| [30] | Zhong L L, Zhou W, Wang H J, Ding S H, Lu Q T, Wen X G, Peng L W, Zhang L X, Lu C M. Chloroplast small heat shock protein HSP21 interacts with plastid nucleoid protein pTAC5 and is essential for chloroplast development in Arabidopsis under heat stress. Plant Cell, 2013,25:2925-2943. |
| [31] |
Bukau B, Weissman J, Horwich A. Molecular chaperones and protein quality control. Cell, 2006,125:443-451.
pmid: 16678092 |
| [32] | Sakuma Y, Maruyama K, Qin F, Osakabe Y, Shinozaki K, Yamaguchi-Shinozaki K. Dual function of an Arabidopsis transcription factor DREB2A in water-stress-responsive and heat-stress- responsive gene expression. Proc Natl Acad Sci USA, 2006,103:18822-18827. |
| [33] | Li X D, Wang X L, Cai Y M, Wu J H, Mo B T, Yu E R. Arabidopsis heat stress transcription factors A2 (HSFA2) and A3 (HSFA3) function in the same heat regulation pathway. Acta Physiol Plant, 2017,39:67-75. |
| [1] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [2] | 王海波, 应静文, 何礼, 叶文宣, 涂卫, 蔡兴奎, 宋波涛, 柳俊. rDNA和端粒重复序列鉴定马铃薯和茄子体细胞杂种染色体丢失和融合[J]. 作物学报, 2022, 48(5): 1273-1278. |
| [3] | 石艳艳, 马志花, 吴春花, 周永瑾, 李荣. 垄作沟覆地膜对旱地马铃薯光合特性及产量形成的影响[J]. 作物学报, 2022, 48(5): 1288-1297. |
| [4] | 周慧文, 丘立杭, 黄杏, 李强, 陈荣发, 范业赓, 罗含敏, 闫海锋, 翁梦苓, 周忠凤, 吴建明. 甘蔗赤霉素氧化酶基因ScGA20ox1的克隆及功能分析[J]. 作物学报, 2022, 48(4): 1017-1026. |
| [5] | 冯亚, 朱熙, 罗红玉, 李世贵, 张宁, 司怀军. 马铃薯StMAPK4响应低温胁迫的功能解析[J]. 作物学报, 2022, 48(4): 896-907. |
| [6] | 张霞, 于卓, 金兴红, 于肖夏, 李景伟, 李佳奇. 马铃薯SSR引物的开发、特征分析及在彩色马铃薯材料中的扩增研究[J]. 作物学报, 2022, 48(4): 920-929. |
| [7] | 谭雪莲, 郭天文, 胡新元, 张平良, 曾骏, 刘晓伟. 黄土高原旱作区马铃薯连作根际土壤微生物群落变化特征[J]. 作物学报, 2022, 48(3): 682-694. |
| [8] | 余慧芳, 张卫娜, 康益晨, 范艳玲, 杨昕宇, 石铭福, 张茹艳, 张俊莲, 秦舒浩. 马铃薯CrRLK1Ls基因家族的鉴定及响应晚疫病菌信号的表达分析[J]. 作物学报, 2022, 48(1): 249-258. |
| [9] | 荐红举, 尚丽娜, 金中辉, 丁艺, 李燕, 王季春, 胡柏耿, Vadim Khassanov, 吕典秋. 马铃薯PIF家族成员鉴定及其对高温胁迫的响应分析[J]. 作物学报, 2022, 48(1): 86-98. |
| [10] | 许德蓉, 孙超, 毕真真, 秦天元, 王一好, 李成举, 范又方, 刘寅笃, 张俊莲, 白江平. 马铃薯StDRO1基因的多态性鉴定及其与根系性状的关联分析[J]. 作物学报, 2022, 48(1): 76-85. |
| [11] | 谢琴琴, 左同鸿, 胡燈科, 刘倩莹, 张以忠, 张贺翠, 曾文艺, 袁崇墨, 朱利泉. 甘蓝自交不亲和相关基因BoPUB9的克隆及表达分析[J]. 作物学报, 2022, 48(1): 108-120. |
| [12] | 苏亚春, 李聪娜, 苏炜华, 尤垂淮, 岑光莉, 张畅, 任永娟, 阙友雄. 甘蔗割手密种类甜蛋白家族鉴定及栽培种同源基因功能分析[J]. 作物学报, 2021, 47(7): 1275-1296. |
| [13] | 李鹏程, 毕真真, 孙超, 秦天元, 梁文君, 王一好, 许德蓉, 刘玉汇, 张俊莲, 白江平. DNA甲基化参与调控马铃薯响应干旱胁迫的关键基因挖掘[J]. 作物学报, 2021, 47(4): 599-612. |
| [14] | 秦天元, 刘玉汇, 孙超, 毕真真, 李安一, 许德蓉, 王一好, 张俊莲, 白江平. 马铃薯StIgt基因家族的鉴定及其对干旱胁迫的响应分析[J]. 作物学报, 2021, 47(4): 780-786. |
| [15] | 杨阳, 李淮琳, 胡利民, 范楚川, 周永明. 白菜型油菜srb多室性状的遗传分析与分子鉴定[J]. 作物学报, 2021, 47(3): 385-393. |
|
||