欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (1): 76-88.doi: 10.3724/SP.J.1006.2024.33007
王丽平( ), 王晓钰, 傅竞也, 王强*()
), 王晓钰, 傅竞也, 王强*()
WANG Li-Ping(), WANG Xiao-Yu, FU Jing-Ye, WANG Qiang*()
摘要:
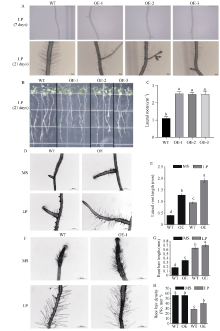
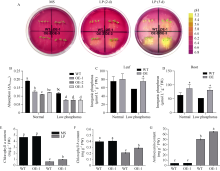
玉米生长发育过程中会遭受干旱、高温、高盐及营养元素匮乏等多种非生物胁迫, 导致其产量和品质下降, 造成严重的农业减产。MYB类转录因子在植物中广泛分布, 参与植物生长发育和环境响应, 筛选及鉴定出具有抗逆功能的MYB类转录因子能为玉米抗逆遗传改良提供理论依据。本研究从玉米干旱处理的材料中克隆了一个R2R3-MYB家族转录因子基因ZmMYB12, 其响应自然干旱、ABA及PEG处理, 基因表达被诱导上调。病毒诱导沉默ZmMYB12后的玉米植株对干旱更加敏感, 活性氧积累更多, 根系更小; 复水后ZmMYB12沉默植株存活率更低, 表明ZmMYB12是干旱正调控因子。进一步构建ZmMYB12稳定过表达拟南芥, 干旱处理后ZmMYB12过表达株系活性氧积累更少, 侧根更多, 抗旱能力增强。同时, 发现ZmMYB12过表达拟南芥在低磷胁迫下侧根数量更多, 根系酸化程度增加, 叶绿素及花青素含量更多, 体内无机磷含量高于野生型, 表明ZmMYB12参与到低磷胁迫过程中, 并且提高了植株对磷元素的吸收及利用率。研究表明ZmMYB12调控干旱抗性并响应低磷胁迫, 为作物抗旱及耐低磷育种提供了一个良好的基因资源。
| [1] |
Campos H, Cooper M, Habben J E, Edmeades G O, Schussler J R. Improving drought tolerance in maize: a view from industry. Field Crops Res, 2004, 90: 19-34.
doi: 10.1016/j.fcr.2004.07.003 |
| [2] |
Esfahanian E, Nejadhashemi A P, Abouali M, Adhikari U, Zhang Z, Daneshvar F, Herman M R. Development and evaluation of a comprehensive drought index. J Environ Manage, 2017, 185: 31-43.
doi: S0301-4797(16)30840-4 pmid: 28029478 |
| [3] |
Gupta A, Rico-Medina A, Caño-Delgado A I. The physiology of plant responses to drought. Science, 2020, 368: 266-269.
doi: 10.1126/science.aaz7614 pmid: 32299946 |
| [4] |
Liu H, Wang X, Wang D, Zou Z, Liang Z. Effect of drought stress on growth and accumulation of active constituents in Salvia miltiorrhiza Bunge. Ind Crop Prod, 2011, 33: 84-88.
doi: 10.1016/j.indcrop.2010.09.006 |
| [5] |
Allen R G, Tresini M, Allen R G, Tresini M. Oxidative stress and gene regulation. Free Radic Biol Med, 2000, 28: 463-499.
doi: 10.1016/S0891-5849(99)00242-7 |
| [6] |
Gong F, Yang L, Tai F, Hu X, Wang W. “Omics” of maize stress response for sustainable food production: opportunities and challenges. OMICS: A J Integr Biol, 2014, 18: 714-732.
doi: 10.1089/omi.2014.0125 |
| [7] |
Pan R, Buitrago S, Feng Z, Abou-Elwafa S F, Xu L, Li C, Zhang W. HvbZIP21, a novel transcription factor from wild barley confers drought tolerance by modulating ROS scavenging. Front Plant Sci, 2022, 13: 878459.
doi: 10.3389/fpls.2022.878459 |
| [8] |
Newton J R. Linked gene ontology categories are novel and differ from associated gene ontology categories for the bipolar disorders. Psychiat Genet, 2007, 17: 29-34.
doi: 10.1097/YPG.0b013e328010f28c |
| [9] |
Ma H Z, Liu C, Li Z X, Ran Q J, Xie G N, Wang B M, Fang S, Chu J F, Zhang J R. ZmbZIP4 contributes to stress resistance in maize by regulating ABA synthesis and root development. Plant Physiol, 2018, 178: 753-770.
doi: 10.1104/pp.18.00436 |
| [10] |
Wang B X, Li L Q, Liu M L, Peng D, Wei A S, Hou B Y, Lei Y H, Li X J. TaFDL2-1A confers drought stress tolerance by promoting ABA biosynthesis, ABA responses, and ROS scavenging in transgenic wheat. Plant J, 2022, 112: 722-737.
doi: 10.1111/tpj.v112.3 |
| [11] |
Zhu J K. Abiotic stress signaling and responses in plants. Cell, 2016, 167: 313-324.
doi: 10.1016/j.cell.2016.08.029 |
| [12] |
Calderón-Vázquez C, Sawers R J, Herrera-Estrella L. Phosphate deprivation in maize: genetics and genomics. Plant Physiol, 2011, 156: 1067-1077.
doi: 10.1104/pp.111.174987 pmid: 21617030 |
| [13] |
Vance C P, Uhde-Stone C, Allan D L. Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytol, 2003, 157: 423-447.
doi: 10.1046/j.1469-8137.2003.00695.x pmid: 33873400 |
| [14] |
Bieleski R L. Phosphate pools, phosphate transport, and phosphate availability. Annu Rev Plant Physiol, 2003, 24: 225-252.
doi: 10.1146/arplant.1973.24.issue-1 |
| [15] |
Usuda H. Phosphate deficiency in maize: V. Mobilization of nitrogen and phosphorus within shoots of young plants and its relationship to senescence. Plant Cell Physiol, 1995, 36: 1041-1049.
doi: 10.1093/oxfordjournals.pcp.a078846 |
| [16] |
Siddiqui M H, Alamri S, Nasir Khan M, Corpas F J, Al-Amri A A, Alsubaie Q D, Ali H M, Kalaji H M, Ahmad P. Melatonin and calcium function synergistically to promote the resilience through ROS metabolism under arsenic-induced stress. J Haz Mat, 2020, 398: 122882.
doi: 10.1016/j.jhazmat.2020.122882 |
| [17] | Bechtold U, Penfold C A, Jenkins D J, Legaie R, Moore J D, Lawson T, Matthews J S, Vialet-Chabrand S R, Baxter L, Subramaniam S, Hickman R, Florance H, Sambles C, Salmon D L, Feil R, Bowden L, Hill C, Baker N R, Lunn J E, Finkenstädt B, Mead A, Buchanan-Wollaston V, Beynon J, Rand D A, Wild D L, Denby K J, Ott S, Smirnoff N, Mullineaux P M. Time-series transcriptomics reveals that AGAMOUS-LIKE22 affects primary metabolism and developmental processes in drought-stressed Arabidopsis. Plant Cell, 2016, 28: 345. |
| [18] |
Bates T R, Lynch J P. Root hairs confer a competitive advantage under low phosphorus availability. Plant Soil, 2001, 236: 243-250.
doi: 10.1023/A:1012791706800 |
| [19] | Devaiah B N, Madhuvanthi R, Karthikeyan A S, Raghothama K G. Phosphate starvation responses and gibberellic acid biosynthesis are regulated by the MYB62 transcription factor in Arabidopsis. Mol Plant, 2009, 2: 43-58. |
| [20] |
Jain A, Poling M D, Smith A P, Nagarajan V K, Lahner B, Meagher R B, Raghothama K G. Variations in the composition of gelling agents affect morphophysiological and molecular responses to deficiencies of phosphate and other nutrients. Plant Physiol, 2009, 150: 1033-1049.
doi: 10.1104/pp.109.136184 pmid: 19386810 |
| [21] |
Wang L S, Li Z, Qian W Q, Guo W L, Gao X, Huang L L, Wang H, Zhu H F, Wu J W, Wang D W, Liu D. The Arabidopsis purple acid phosphatase AtPAP10 is predominantly associated with the root surface and plays an important role in plant tolerance to phosphate limitation. Plant Physiol, 2011, 157: 1283-1299.
doi: 10.1104/pp.111.183723 |
| [22] |
Zhang J Y, Zhou X, Xu Y, Yao M L, Xie F B, Gai J Y, Li Y, Yang S P. Soybean SPX1 is an important component of the response to phosphate deficiency for phosphorus homeostasis. Plant Sci, 2016, 248: 82-91.
doi: 10.1016/j.plantsci.2016.04.010 pmid: 27181950 |
| [23] |
Martin C, Paz-Ares J. MYB transcription factors in plants. Trends Genet, 1997, 13: 67-73.
doi: 10.1016/s0168-9525(96)10049-4 pmid: 9055608 |
| [24] |
Fang Q, Wang X Q, Wang H Y, Tang X W, Liu C, Yin H, Ye S L, Jiang Y Z, Duan Y J, Luo K M. The poplar R2R3 MYB transcription factor PtrMYB94 coordinates with abscisic acid signaling to improve drought tolerance in plants. Tree Physiol, 2020, 40: 46-59.
doi: 10.1093/treephys/tpz113 pmid: 31728530 |
| [25] | Gao F, Zhou J, Deng R Y, Zhao H X, Li C L, Chen H, Suzuki T, Park S U, Wu Q.Overexpression of a tartary buckwheat R2R3-MYB transcription factor gene, FtMYB9, enhances tolerance to drought and salt stresses in transgenic Arabidopsis. Plant Physiol, 2017, 214: 81-90. |
| [26] |
Shukla P S, Gupta K, Agarwal P, Jha B, Agarwal P K. Overexpression of a novel SbMYB15 from Salicornia brachiata confers salinity and dehydration tolerance by reduced oxidative damage and improved photosynthesis in transgenic tobacco. Planta, 2015, 242: 1291-308.
doi: 10.1007/s00425-015-2366-5 pmid: 26202734 |
| [27] |
Zheng X W, Liu C, Qiao L, Zhao J J, Han R, Wang X L, Ge C, Zhang W Y, Zhang S W, Qiao L Y, Zheng J, Hao C Y. The MYB transcription factor TaPHR3-A1 is involved in phosphate signaling and governs yield-related traits in bread wheat. J Exp Bot, 2020, 71: 5808-5822.
doi: 10.1093/jxb/eraa355 pmid: 32725154 |
| [28] |
Gulzar F, Fu J Y, Zhu C Y, Yan J, Li X L, Meraj T A, Shen Q Q, Hassan B, Wang Q. Maize WRKY transcription factor ZmWRKY79 positively regulates drought tolerance through elevating ABA biosynthesis. Int J Mol Sci, 2021, 22: 10080.
doi: 10.3390/ijms221810080 |
| [29] | Pu Q Y, Liang J, Shen Q Q, Fu J Y, Pu Z E, Liu J, Wang X G, Wang Q. A wheat β-patchoulene synthase confers resistance against herbivory in transgenic Arabidopsis. Genes, 2019, 10: 441. |
| [30] |
Wu J D, Jiang Y L, Liang Y N, Chen L, Chen W J, Cheng B J. Expression of the maize MYB transcription factor ZmMYB3R enhances drought and salt stress tolerance in transgenic plants. Plant Physiol Bioch, 2019, 137: 179-188.
doi: 10.1016/j.plaphy.2019.02.010 |
| [31] |
Du H, Feng B R, Yang S S, Huang Y B, Tang Y X, Wu K. The R2R3-MYB transcription factor gene family in Maize. PLoS One, 2012, 7: e37463.
doi: 10.1371/journal.pone.0037463 |
| [32] |
Castorina G, Domergue F, Chiara M, Zilio M, Persico M, Ricciardi V, Horner D S, Consonni G. ZmFDL1/MYB94 drought- responsive regulates cuticle biosynthesis and cuticle-dependent leaf permeability. Plant Physiol, 2020, 184: 266-282.
doi: 10.1104/pp.20.00322 pmid: 32665334 |
| [33] |
Ogawa M, Hanada A, Yamauchi Y, Kuwahara A, Kamiya Y, Yamaguchi S. Gibberellin biosynthesis and response during Arabidopsis seed germination. Plant Cell, 2003, 15: 1591-1604.
doi: 10.1105/tpc.011650 |
| [34] | Zhong C M, Patra B, Tang Y, Li X K, Yuan L, Wang X J. A transcriptional hub integrating gibberellin-brassinosteroid signals to promote seed germination in Arabidopsis. J Exp Bot, 2021, 72: 4708-4720. |
| [35] |
Sano N, Marion-Poll A. ABA metabolism and homeostasis in seed dormancy and germination. Int J Mol Sci, 2021, 22: 5069.
doi: 10.3390/ijms22105069 |
| [36] |
Wang X P, Niu Y L, Zheng Y. Multiple Functions of MYB transcription factors in abiotic stress responses. Int J Mol Sci, 2021, 22: 6125.
doi: 10.3390/ijms22116125 |
| [37] | Chen K, Li G J, A Bressan R, Song C P, Zhu J K. Abscisic acid dynamics, signaling, and functions in plants. J Integr Plant Biol, 2020, 62: 27-56. |
| [38] |
Xu W, Tang W, Wang C, Ge L, Sun J, Qi X, He Z, Zhou Y, Chen J, Xu Z, Ma Y Z, Chen M. SiMYB56 confers drought stress tolerance in transgenic rice by regulating lignin biosynthesis and ABA signaling pathway. Front Plant Sci, 2020, 11: 785.
doi: 10.3389/fpls.2020.00785 pmid: 32625221 |
| [39] |
Valdés-López O, Arenas-Huertero C, Ramírez M, Girard L, Sánchez F, Vance C P, Luis Reyes J, Hernández G. Essential role of MYB transcription factor: PvPHR1 and microRNA: PvmiR399 in phosphorus-deficiency signalling in common bean roots. Plant Cell Environ, 2008, 31: 1834-1843.
doi: 10.1111/pce.2008.31.issue-12 |
| [40] |
Val-Torregrosa B, Bundó M, Martín-Cardoso H, Bach-Pages M, Chiou T J, Flors V, Segundo B S. Phosphate-induced resistance to pathogen infection in Arabidopsis. Plant J, 2022, 110: 452-469.
doi: 10.1111/tpj.v110.2 |
| [41] |
Tariq A, Pan K, Olatunji O A, Graciano C, Li Z, Sun F Sun X, Song D, Chen W, Zhang A, Wu X, Zhang L, Mingrui D, Xiong Q, Liu C. Phosphorous application improves drought tolerance of Phoebe zhennan. Front Plant Sci, 2017, 8: 1561.
doi: 10.3389/fpls.2017.01561 pmid: 28955356 |
| [42] |
Begum N, Ahanger M A, Zhang L X. AMF inoculation and phosphorus supplementation alleviates drought induced growth and photosynthetic decline in Nicotiana tabacum by up-regulating antioxidant metabolism and osmolyte accumulation. Environ Exp Bot, 2020, 176: 104088.
doi: 10.1016/j.envexpbot.2020.104088 |
| [43] |
Hansel F D, Amado T J C, Ruiz Diaz D A, Rosso L H M, Schorr M. Phosphorus fertilizer placement and tillage affect soybean root growth and drought tolerance. Agron J, 2017, 109: 2936-2944.
doi: 10.2134/agronj2017.04.0202 |
| [1] | 吴昊, 张瑛, 王琛, 顾汉柱, 周天阳, 张伟杨, 顾骏飞, 刘立军, 杨建昌, 张耗. 栽培优化对长江下游水稻灌浆期根系特征和稻米淀粉特性的影响[J]. 作物学报, 2024, 50(2): 478-492. |
| [2] | 毛燕, 郑名敏, 牟成香, 谢吴兵, 唐琦. 渗透胁迫下玉米自然反义转录本cis-NATZmNAC48启动子的功能分析[J]. 作物学报, 2024, 50(2): 354-362. |
| [3] | 马娟, 曹言勇 . 玉米杂交群体产量性状及其特殊配合力全基因组关联分析[J]. 作物学报, 2024, 50(2): 363-372. |
| [4] | 杨静蕾, 吴冰杰, 王安洲, 肖英杰. 基于多维组学数据的玉米农艺和品质性状预测研究[J]. 作物学报, 2024, 50(2): 373-382. |
| [5] | 朱晓亚, 张强强, 赵鹏, 刘明, 王静, 靳容, 于永超, 唐忠厚. 叶面喷施丹参碳点缓解甘薯低磷胁迫的转录组与代谢组学分析[J]. 作物学报, 2024, 50(2): 383-393. |
| [6] | 吴宇, 刘磊, 崔克辉, 齐晓丽, 黄见良, 彭少兵. 低氮条件下超级杂交稻苗期根系特征的变化及与其高氮素积累的关系[J]. 作物学报, 2024, 50(2): 414-424. |
| [7] | 徐冉, 杨文叶, 朱均林, 陈松, 徐春梅, 刘元辉, 章秀福, 王丹英, 褚光. 不同灌溉模式对籼粳杂交稻甬优1540产量与水分利用效率的影响[J]. 作物学报, 2024, 50(2): 425-439. |
| [8] | 李艳, 方宇辉, 王永霞, 彭超军, 华夏, 齐学礼, 胡琳, 许为钢. 不同磷胁迫处理转OsPHR2小麦的转录组学分析[J]. 作物学报, 2024, 50(2): 340-353. |
| [9] | 杨晨曦, 周文期, 周香艳, 刘忠祥, 周玉乾, 刘芥杉, 杨彦忠, 何海军, 王晓娟, 连晓荣, 李永生. 控制玉米株高基因PHR1的基因克隆[J]. 作物学报, 2024, 50(1): 55-66. |
| [10] | 岳润清, 李文兰, 孟昭东. 转基因抗虫耐除草剂玉米自交系LG11的获得及抗性分析[J]. 作物学报, 2024, 50(1): 89-99. |
| [11] | 宋旭东, 朱广龙, 张舒钰, 章慧敏, 周广飞, 张振良, 冒宇翔, 陆虎华, 陈国清, 石明亮, 薛林, 周桂生, 郝德荣. 长江中下游地区糯玉米花期耐热性鉴定及评价指标筛选[J]. 作物学报, 2024, 50(1): 172-186. |
| [12] | 杨立达, 任俊波, 彭新月, 杨雪丽, 罗凯, 陈平, 袁晓婷, 蒲甜, 雍太文, 杨文钰. 施氮与种间距离下大豆/玉米带状套作作物生长特性及其对产量形成的影响[J]. 作物学报, 2024, 50(1): 251-264. |
| [13] | 谭志新, 谢留伟, 李洪戈, 李芳军, 田晓莉, 李召虎. 基于AHP-隶属函数法的棉花子叶期耐低钾能力鉴定[J]. 作物学报, 2024, 50(1): 199-208. |
| [14] | 艾蓉, 张春, 悦曼芳, 邹华文, 吴忠义. 玉米转录因子ZmEREB211对非生物逆境胁迫的应答[J]. 作物学报, 2023, 49(9): 2433-2445. |
| [15] | 黄钰杰, 张啸天, 陈会丽, 王宏伟, 丁双成. 玉米ZmC2s基因家族鉴定及ZmC2-15耐热功能分析[J]. 作物学报, 2023, 49(9): 2331-2343. |
|