欢迎访问作物学报,今天是
作物学报 ›› 2025, Vol. 51 ›› Issue (4): 982-991.doi: 10.3724/SP.J.1006.2025.44147
郭绪虎1,*( ), 李灵芝2, 李凤1, 马博岩1, 贾晓宇1
), 李灵芝2, 李凤1, 马博岩1, 贾晓宇1
GUO Xu-Hu1,*(), LI Ling-Zhi2, LI Feng1, MA Bo-Yan1, JIA Xiao-Yu1
摘要:
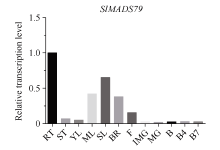
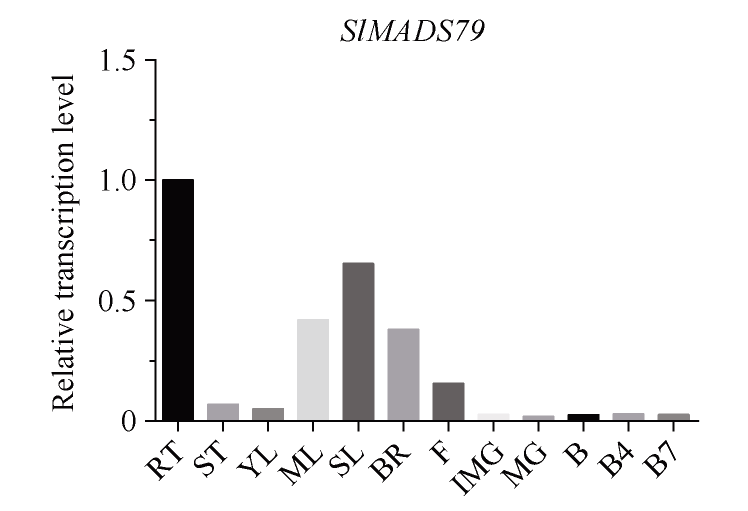
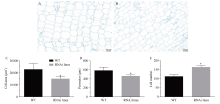
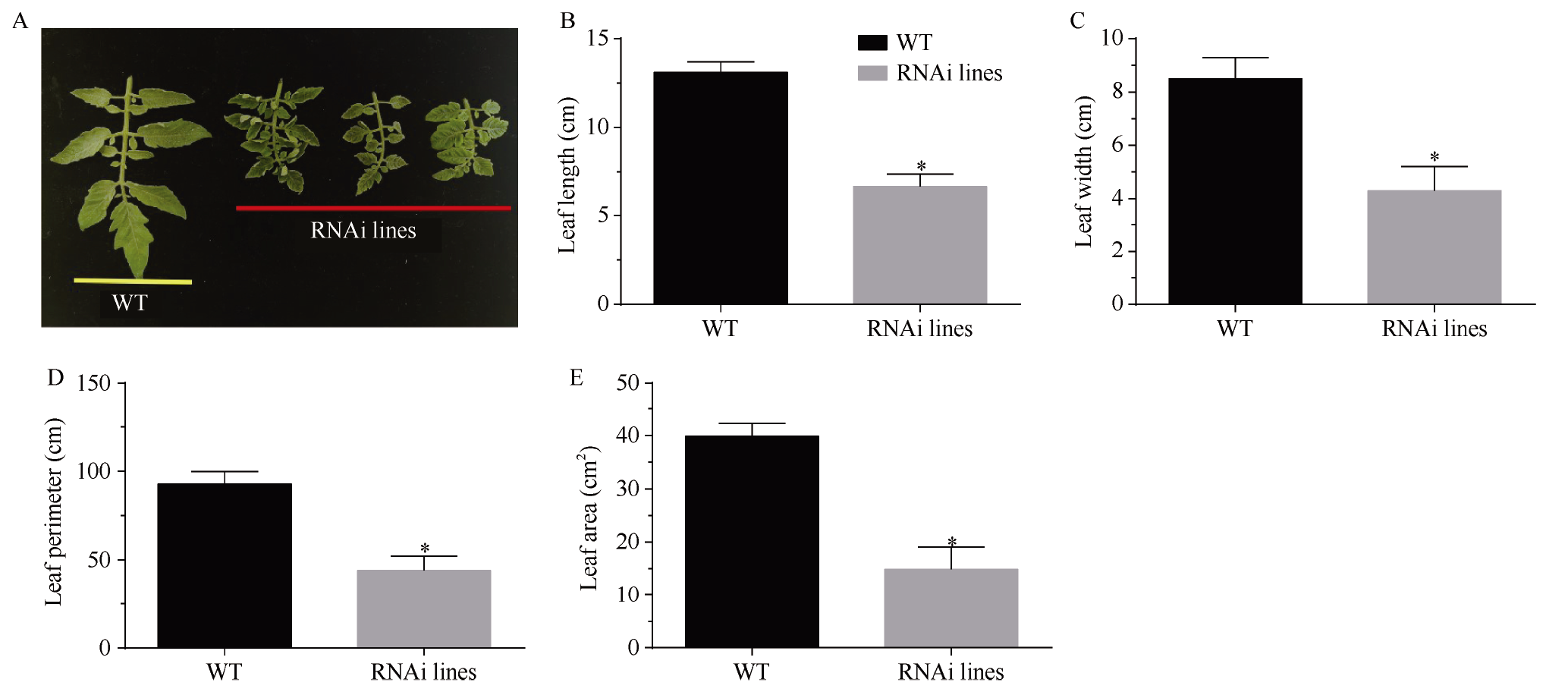
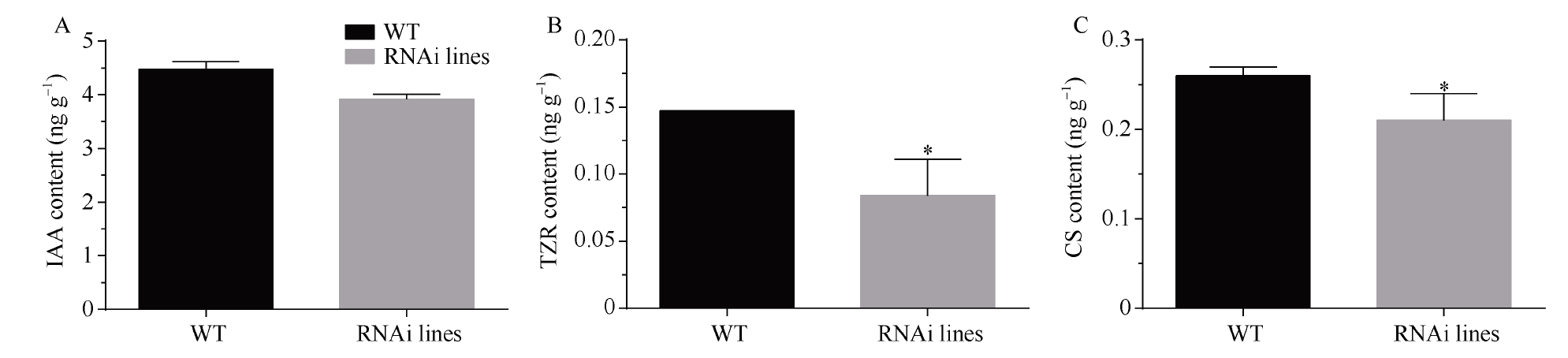
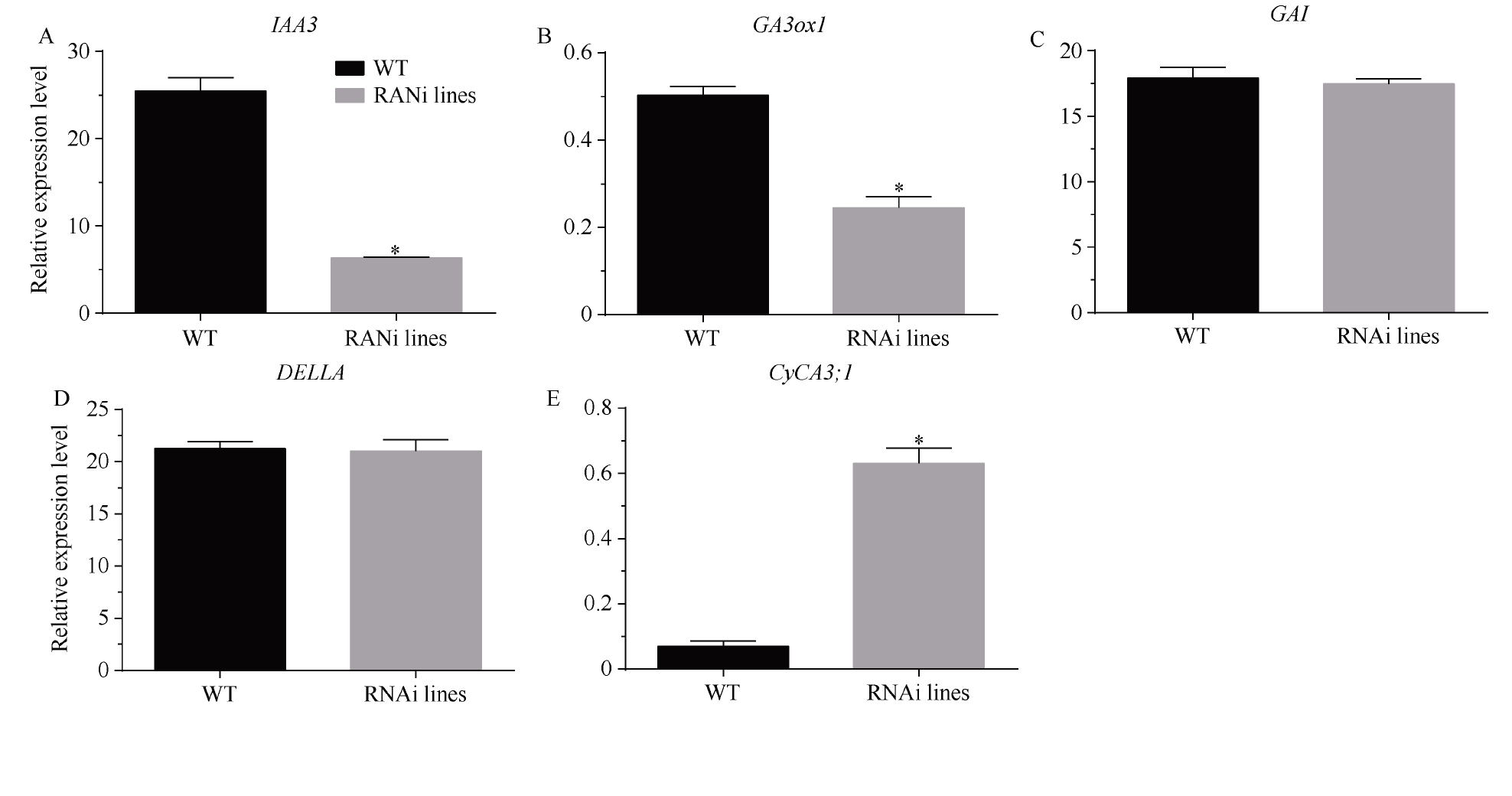
MADS-box基因在植物生长发育全过程中具有重要调控功能。目前, 关于番茄II型MADS-box基因的功能研究较多, 而I型MADS-box基因却鲜有报道。本研究克隆了I型MADS-box基因SlMADS79, 该基因在番茄根、叶和侧芽中表达水平相对较高, 推测其可能参与番茄营养器官生长调控。本研究以经典番茄栽培品种Ailsa Craig (AC++)为背景材料, 采用RNA干扰技术沉默SlMADS79基因。与野生型相比, SlMADS79基因沉默株系顶端优势度降低、植株矮小; 叶片的长度、宽度、周长以及面积均小于野生型植株; 根系总长度、总表面积、总投影面积、根系体积、分叉数、根尖数均显著减少。解剖学研究表明, 沉默植株茎秆的纵切面细胞较小, 然而其平均数量明显增加。在激素水平上, SlMADS79沉默株系中IAA (吲哚-3-乙酸)、TZR (转玉米素核糖苷)以及CS (栗木甾酮)含量降低。在分子水平上, 生长素应答基因IAA3和赤霉素合成基因GA3ox1在SlMADS79沉默株系中显著下调, 而细胞周期基因CyCA3;1在沉默株系中显著上调。本研究从形态、解剖、激素和分子水平上进一步解析了SlMADS79基因的生物学功能, 丰富了番茄I型MADS-box基因家族的研究, 为番茄株型调控研究提供了可靠的理论依据。
| [1] |
Alvarez-Buylla E R, Pelaz S, Liljegren S J, Gold S E, Burgeff C, Ditta G S, Ribas de Pouplana L, Martínez-Castilla L, Yanofsky M F. An ancestral MADS-box gene duplication occurred before the divergence of plants and animals. Proc Natl Acad Sci USA, 2000, 97: 5328-5333.
pmid: 10805792 |
| [2] |
Smaczniak C, Immink R G H, Angenent G C, Kaufmann K. Developmental and evolutionary diversity of plant MADS-domain factors: insights from recent studies. Development, 2012, 139: 3081-3098.
doi: 10.1242/dev.074674 pmid: 22872082 |
| [3] |
Henschel K, Kofuji R, Hasebe M, Saedler H, Münster T, Theissen G. Two ancient classes of MIKC-type MADS-box genes are present in the moss Physcomitrella patens. Mol Biol Evol, 2002, 19: 801-814.
pmid: 12032236 |
| [4] | Yu L H, Miao Z Q, Qi G F, Wu J, Cai X T, Mao J L, Xiang C B. MADS-box transcription factor AGL21 regulates lateral root development and responds to multiple external and physiological signals. Mol Plant, 2014, 7: 1653-1669. |
| [5] |
Teper-Bamnolker P, Samach A. The flowering integrator FT regulates SEPALLATA3 and FRUITFULL accumulation in Arabidopsis leaves. Plant Cell, 2005, 17: 2661-2675.
doi: 10.1105/tpc.105.035766 pmid: 16155177 |
| [6] | Zhou Y, Hu L F, Song J B, Jiang L W, Liu S Q. Isolation and characterization of a MADS-box gene in cucumber (Cucumis sativus L.) that affects flowering time and leaf morphology in transgenic Arabidopsis. Biotechnol Biotec Equip, 2019, 33: 54-63. |
| [7] | Xing M Y, Li H L, Liu G S, Zhu B Z, Zhu H L, Grierson D, Luo Y B, Fu D Q. A MADS-box transcription factor, SlMADS1, interacts with SlMACROCALYX to regulate tomato sepal growth. Plant Sci, 2022, 322: 111366. |
| [8] | Guo X H, Chen G P, Naeem M, Yu X H, Tang B Y, Li A Z, Hu Z L. The MADS-box gene SlMBP11 regulates plant architecture and affects reproductive development in tomato plants. Plant Sci, 2017, 258: 90-101. |
| [9] | Li A Z, Chen G P, Yu X H, Zhu Z G, Zhang L C, Zhou S G, Hu Z L. The tomato MADS-box gene SlMBP9 negatively regulates lateral root formation and apical dominance by reducing auxin biosynthesis and transport. Plant Cell Rep, 2019, 38: 951-963. |
| [10] | Yin W C, Yu X H, Chen G P, Tang B Y, Wang Y S, Liao C G, Zhang Y J, Hu Z L. Suppression of SlMBP15 inhibits plant vegetative growth and delays fruit ripening in tomato. Front Plant Sci, 2018, 9: 938. |
| [11] | Wang Y S, Guo P Y, Zhang J L, Xie Q L, Shen H, Hu Z L, Chen G P. Overexpression of the MADS-box gene SIMBP21 alters leaf morphology and affects reproductive development in tomato. J Integr Agric, 2021, 20: 3170-3185. |
| [12] | Li F F, Chen X Y, Zhou S E, Xie Q L, Wang Y S, Xiang X X, Hu Z L, Chen G P. Overexpression of SlMBP22 in tomato affects plant growth and enhances tolerance to drought stress. Plant Sci, 2020, 301: 110672. |
| [13] | Kojima M, Kamada-Nobusada T, Komatsu H, Takei K, Kuroha T, Mizutani M, Ashikari M, Ueguchi-Tanaka M, Matsuoka M, Suzuki K, Sakakibara H. Highly sensitive and high-throughput analysis of plant hormones using MS-probe modification and liquid chromatography-tandem mass spectrometry: an application for hormone profiling in Oryza sativa. Plant Cell Physiol, 2009, 50: 1201-1214. |
| [14] |
Pan X Q, Welti R, Wang X M. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat Protoc, 2010, 5: 986-992.
doi: 10.1038/nprot.2010.37 pmid: 20448544 |
| [15] |
Expósito-Rodríguez M, Borges A A, Borges-Pérez A, Pérez J A. Selection of internal control genes for quantitative real-time RT-PCR studies during tomato development process. BMC Plant Biol, 2008, 8: 131.
doi: 10.1186/1471-2229-8-131 pmid: 19102748 |
| [16] |
Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔСT method. Methods, 2001, 25: 402-408.
doi: 10.1006/meth.2001.1262 pmid: 11846609 |
| [17] | Pelaz S, Ditta G S, Baumann E, Wisman E, Yanofsky M F. B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature, 2000, 405: 200-203. |
| [18] |
Michaels S D, Ditta G, Gustafson-Brown C, Pelaz S, Yanofsky M, Amasino R M. AGL24 acts as a promoter of flowering in Arabidopsis and is positively regulated by vernalization. Plant J, 2003, 33: 867-874.
doi: 10.1046/j.1365-313x.2003.01671.x pmid: 12609028 |
| [19] |
Dong T T, Hu Z L, Deng L, Wang Y, Zhu M K, Zhang J L, Chen G P. A tomato MADS-box transcription factor, SlMADS1, acts as a negative regulator of fruit ripening. Plant Physiol, 2013, 163: 1026-1036.
doi: 10.1104/pp.113.224436 pmid: 24006286 |
| [20] | Wang Y S, Zhang J L, Hu Z L, Guo X H, Tian S B, Chen G P. Genome-wide analysis of the MADS-box transcription factor family in Solanum lycopersicum. Int J Mol Sci, 2019, 20: 2961. |
| [21] |
李艳大, 朱相成, 汤亮, 曹卫星, 朱艳. 基于株型的水稻冠层光合生产模拟. 作物学报, 2011, 37: 868-875.
doi: 10.3724/SP.J.1006.2011.00868 |
| Li Y D, Zhu X C, Tang L, Cao W X, Zhu Y. Simulation of canopy photosynthetic production based on plant type in rice. Acta Agron Sin, 2011, 37: 868-875 (in Chinese with English abstract). | |
| [22] |
许娜, 徐铨, 徐正进, 陈温福. 水稻株型生理生态与遗传基础研究进展. 作物学报, 2023, 49: 1735-1746.
doi: 10.3724/SP.J.1006.2023.22050 |
|
Xu N, Xu Q, Xu Z J, Chen W F. Research progress on physiological ecology and genetic basis of rice plant architecture. Acta Agron Sin, 2023, 49: 1735-1746 (in Chinese with English abstract).
doi: 10.3724/SP.J.1006.2023.22050 |
|
| [23] | Li A Z, Chen G P, Wang Y S, Liang H L, Hu Z L. Silencing of the MADS-box gene SlMADS83 enhances adventitious root formation in tomato plants. J Plant Growth Regul, 2020, 39: 941-953. |
| [24] |
Kazan K, Manners J M. Linking development to defense: auxin in plant-pathogen interactions. Trends Plant Sci, 2009, 14: 373-382.
doi: 10.1016/j.tplants.2009.04.005 pmid: 19559643 |
| [25] | Pang Y, Zang X Y, Pang F T, Zhou T H, Tian F Z. Changes of CTK and few nitrogen index during development of flower and fruit in Zhanhua jujube. J North China Agric, 2017, 5: 101. |
| [26] | Grove M D, Spencer G F, Rohwedder W K, Mandava N, Worley J F, Warthen J D, Steffens G L, Flippen-Anderson J L, Cook J C. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nature, 1979, 281: 216-217. |
| [27] |
Clouse S D, Langford M, McMorris T C. A brassinosteroid- insensitive mutant in Arabidopsis thaliana exhibits multiple defects in growth and development. Plant Physiol, 1996, 111: 671-678.
doi: 10.1104/pp.111.3.671 pmid: 8754677 |
| [28] |
Reed J W. Roles and activities of Aux/IAA proteins in Arabidopsis. Trends Plant Sci, 2001, 6: 420-425.
doi: 10.1016/s1360-1385(01)02042-8 pmid: 11544131 |
| [29] |
Wang H, Jones B, Li Z G, Frasse P, Delalande C, Regad F, Chaabouni S, Latché A, Pech J C, Bouzayen M. The tomato Aux/IAA transcription factor IAA9 is involved in fruit development and leaf morphogenesis. Plant Cell, 2005, 17: 2676-2692.
doi: 10.1105/tpc.105.033415 pmid: 16126837 |
| [30] |
Chaabouni S, Jones B, Delalande C, Wang H, Li Z G, Mila I, Frasse P, Latché A, Pech J C, Bouzayen M. Sl-IAA3, a tomato Aux/IAA at the crossroads of auxin and ethylene signalling involved in differential growth. J Exp Bot, 2009, 60: 1349-1362.
doi: 10.1093/jxb/erp009 pmid: 19213814 |
| [31] |
Hooley R. Gibberellins: perception, transduction and responses. Plant Mol Biol, 1994, 26: 1529-1555.
pmid: 7858203 |
| [32] |
Swain S M, Olszewski N E. Genetic analysis of gibberellin signal transduction. Plant Physiol, 1996, 112: 11-17.
pmid: 12226370 |
| [33] |
Yamaguchi S. Gibberellin metabolism and its regulation. Annu Rev Plant Biol, 2008, 59: 225-251.
doi: 10.1146/annurev.arplant.59.032607.092804 pmid: 18173378 |
| [34] |
Vogler H, Caderas D, Mandel T, Kuhlemeier C. Domains of expansin gene expression define growth regions in the shoot apex of tomato. Plant Mol Biol, 2003, 53: 267-272.
doi: 10.1023/b:plan.0000006999.48516.be pmid: 14750517 |
| [35] |
Zhang T Y, Wang X, Lu Y E, Cai X F, Ye Z B, Zhang J H. Genome-wide analysis of the cyclin gene family in tomato. Int J Mol Sci, 2013, 15: 120-140.
doi: 10.3390/ijms15010120 pmid: 24366066 |
| [36] | Peng J, Carol P, Richards D E, King K E, Cowling R J, Murphy G P, Harberd N P. The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses. Genes Dev, 1997, 11: 3194-3205. |
| [1] | 徐建霞, 丁延庆, 曹宁, 程斌, 高旭, 李文贞, 张立异. 中国高粱株高和节间数全基因组关联分析及候选基因预测[J]. 作物学报, 2025, 51(3): 568-585. |
| [2] | 张辰煜, 葛军勇, 褚俊聪, 王星宇, 赵宝平, 杨亚东, 臧华栋, 曾昭海. 燕麦红芸豆带状间作的产量效应及根系形态与土壤酶活性[J]. 作物学报, 2025, 51(2): 459-469. |
| [3] | 郭思语, 赵克勇, 代正罡, 邹华文, 吴忠义, 张春. 玉米N-乙酰转移酶ZmNAT1基因响应非生物胁迫的功能分析[J]. 作物学报, 2024, 50(8): 2001-2013. |
| [4] | 刘宸铭, 赵克勇, 悦曼芳, 赵延明, 吴忠义, 张春. 玉米转录因子ZmEREB180调控根系生长发育及耐逆的功能研究[J]. 作物学报, 2024, 50(8): 1920-1933. |
| [5] | 韩笑晨, 张贵芹, 王亚辉, 任昊, 王洪章, 刘国利, 林佃旭, 王子强, 张吉旺, 赵斌, 任佰朝, 刘鹏. 土壤调理剂对滨海盐碱地土壤盐分含量及夏玉米产量的影响[J]. 作物学报, 2024, 50(7): 1776-1786. |
| [6] | 唐清芸, 杨晶晶, 赵蕾, 宋志文, 王国栋, 李玉祥. 施氮量对滴灌水稻根系形态构型和分形特征的影响[J]. 作物学报, 2024, 50(6): 1540-1553. |
| [7] | 周香玉, 徐劲松, 谢伶俐, 许本波, 张学昆. 甘蓝型油菜苗期响应渍害胁迫的生理调控机制[J]. 作物学报, 2024, 50(4): 1015-1029. |
| [8] | 刘薇, 王玉斌, 李伟, 张礼凤, 徐冉, 王彩洁, 张彦威. 过量表达大豆异丙基苹果酸脱氢酶基因GmIPMDH促进植株开花和生长[J]. 作物学报, 2024, 50(3): 613-622. |
| [9] | 吴昊, 张瑛, 王琛, 顾汉柱, 周天阳, 张伟杨, 顾骏飞, 刘立军, 杨建昌, 张耗. 栽培优化对长江下游水稻灌浆期根系特征和稻米淀粉特性的影响[J]. 作物学报, 2024, 50(2): 478-492. |
| [10] | 刁现民, 王立伟, 智慧, 张俊, 李顺国, 程汝宏. 谷子中矮秆资源创制、遗传解析和育种利用[J]. 作物学报, 2024, 50(2): 265-279. |
| [11] | 吴宇, 刘磊, 崔克辉, 齐晓丽, 黄见良, 彭少兵. 低氮条件下超级杂交稻苗期根系特征的变化及与其高氮素积累的关系[J]. 作物学报, 2024, 50(2): 414-424. |
| [12] | 徐冉, 杨文叶, 朱均林, 陈松, 徐春梅, 刘元辉, 章秀福, 王丹英, 褚光. 不同灌溉模式对籼粳杂交稻甬优1540产量与水分利用效率的影响[J]. 作物学报, 2024, 50(2): 425-439. |
| [13] | 王成, 马杨明, 王春雨, 李志欣, 罗健升, 彭政岚, 刘儒宏基, 黄兴海, 曹云, 彭政菠, 马均. 种植方式与施氮量对杂交籼稻养分吸收特性及根系活力的影响[J]. 作物学报, 2024, 50(12): 3069-3082. |
| [14] | 戎宇轩, 惠留洋, 王沛琦, 孙思敏, 张献龙, 袁道军, 杨细燕. 陆地棉CLE基因家族的鉴定及GhCLE13参与调控棉花抗旱性的功能分析[J]. 作物学报, 2024, 50(12): 2925-2939. |
| [15] | 阳世杰, 王华智, 潘怡敏, 黄蕊, 侯森, 秦慧彬, 穆志新, 王海岗. 山西谷子种质资源株高全基因组关联分析[J]. 作物学报, 2024, 50(12): 2984-2997. |
|
||