欢迎访问作物学报,今天是
作物学报 ›› 2018, Vol. 44 ›› Issue (9): 1290-1300.doi: 10.3724/SP.J.1006.2018.01290
李红丹1,2,闫蕾1,2,孙蕾1,2,樊晓聪1,3,陈士瞻1,3,张燕1,3,郭林1,游光霞1,李庄1,2,杨宗举1,2,苏亮1,*,杨建平1,3,*
Hong-Dan LI1,2,Lei YAN1,2,Lei SUN1,2,Xiao-Cong FAN1,3,Shi-Zhan CHEN1,3,Yan ZHANG1,3,Lin GUO1,Guang-Xia YOU1,Zhuang LI1,2,Zong-Ju YANG1,2,Liang SU1,*,Jian-Ping YANG1,3,*
摘要:
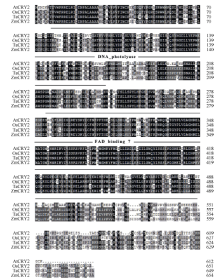
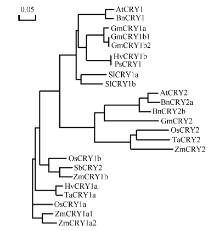
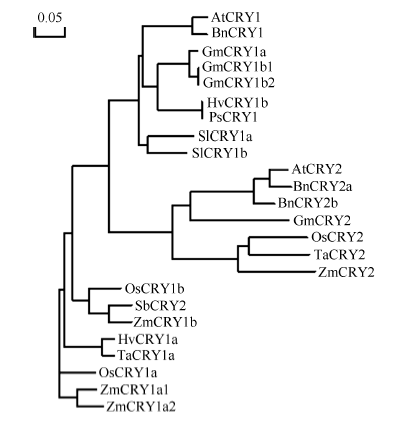
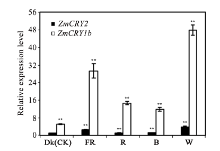
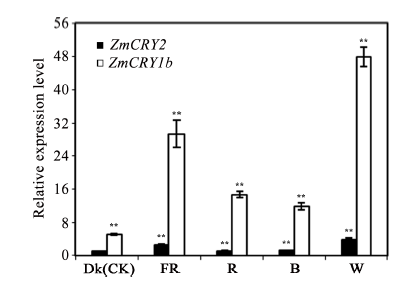
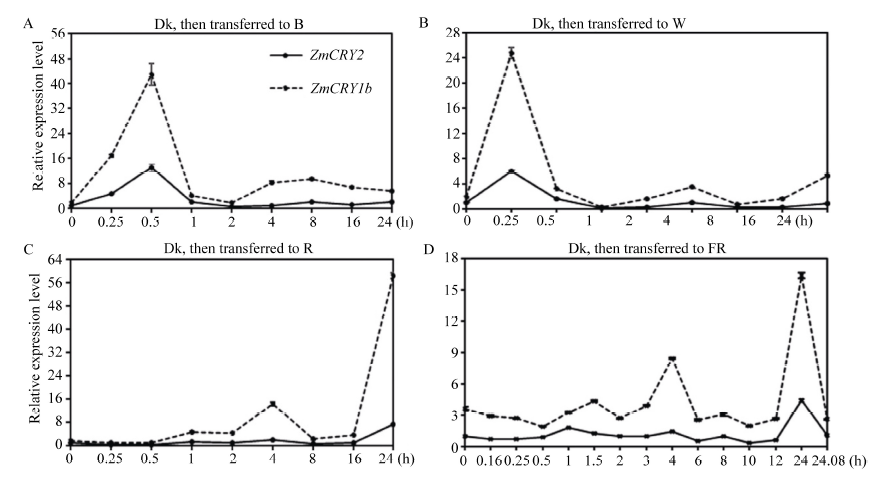
玉米株高、开花期、产量、品质等性状与环境中的光密切相关。隐花色素是一类蓝光和近紫外光的受体, 主要参与植物的光形态建成及动、植物的生物钟调控。通过研究玉米隐花色素基因对不同光处理的表达模式, 可为进一步研究其对玉米光形态建成的作用奠定基础。本研究采用RT-PCR技术克隆了玉米ZmCRY1b和ZmCRY2基因; 利用生物信息学相关网站和软件对其编码蛋白的结构域及氨基酸进行了系统发育分析; 利用qRT-PCR分析了玉米自交系B73中ZmCRY1b和ZmCRY2基因在不同组织、以及响应不同光质及长日照和短日照处理的转录丰度。研究发现, 玉米与拟南芥、水稻和小麦的CRY蛋白有相同的结构域及较高的氨基酸序列的一致性, 表明它们具有相似的功能。ZmCRY1b和ZmCRY2基因主要在玉米的叶片中表达; 二者能迅速响应各种持续光质、黑暗到不同光质转换及长日照和短日照处理, 且ZmCRY1b在各种处理下的转录丰度均高于ZmCRY2, 可能暗示ZmCRY1b在玉米中功能更强。以上研究结果表明, ZmCRY1b和ZmCRY2基因均能有效地响应各种光质和光周期处理, 并在玉米的光形态建成中发挥重要作用。本研究为进一步探明ZmCRY1b和ZmCRY2基因的功能及其在玉米品种改良中的应用提供了研究基础。
| [1] | 詹克慧, 李志勇, 侯佩, 习雨琳, 肖阳, 孟凡华, 杨建平 . 利用修饰光敏色素信号途径进行品种改良的可行性. 中国农业科学, 2012,45:3249-3255 |
| Zhan K H, Li Z Y, Hou P, Xi Y L, Xiao Y, Meng F H, Yang J P . A new strategy for crop improvement through modification of phytochrome signaling pathways. Sci Agric Sin, 2012,45:3249-3255 (in Chinese with English abstract) | |
| [2] | Weller J L, Perrotta G , Schreuder M E, van Tuinen A, Koornneef M, Giuliano G, Kendrick R E . Genetic dissection of blue-light sensing in tomato using mutants deficient in cryptochrome 1 and phytochromes A, B1 and B2. Plant J, 2001,25:427-440 |
| [3] | Giliberto L, Perrotta G, Pallara P, Weller J L, Fraser P D, Bramley P M, Fiore A, Tavazza M, Giuliano G . Manipulation of the blue light photoreceptor cryptochrome 2 in tomato affects vegetative development, flowering time and fruit antioxidant content. Plant Physiol, 2005,137:199-208 |
| [4] |
Platten J D, Foo E, Elliott R C, Hecht V, Reid J B, Weller J L . Cryptochrome 1 contributes to blue-light sensing in pea. Plant Physiol, 2005,139:1472-1482
doi: 10.1104/pp.105.067462 pmid: 16244154 |
| [5] | Sharma P, Chatterjee M, Burman N, Khurana J P . Cryptochrome 1 regulates growth and development in Brassica through alteration in the expression of genes involved in light, phytohormone and stress signalling. Plant Cell Environ, 2014,37:961-977 |
| [6] |
Yang Z H, Liu B B, Su J, Liao J K, Lin C T, Oka Y . Cryptochromes orchestrate transcription regulation of diverse blue light responses in plants. Photochem Photobiol, 2017,93:112-127
doi: 10.1111/php.12663 pmid: 27861972 |
| [7] | Sadanandom A, Ádám É, Orosa B, Viczián A, Klose C, Zhang C, Josse E, Kozma-Bognár L, Nagy F . SUMOylation of phytochrome-B negatively regulates light-induced signaling in Arabidopsis thaliana. Proc Natl Acad Sci USA, 2015,112:11108-11113 |
| [8] | Liu B, Yang Z H, Adam Gomez, Liu B, Lin C T, Oka Y . Signaling mechanisms of plant cryptochromes in Arabidopsis thaliana. J Plant Res, 2016,129:137-148 |
| [9] |
Yuan S, Zhang Z W, Zheng C, Zhao Z Y, Wang Y, Feng L Y, Niu G Q, Wang C Q, Wang J H, Feng H, Xu F, Bao F, Hua Y, Cao Y, Ma L G, Wang H Y, Kong D D, Xiao W, Lin H H, He Y K . Arabidopsis cryptochrome 1 functions in nitrogen regulation of flowering. Proc Natl Acad Sci USA, 2016,113:7661-7666
doi: 10.1073/pnas.1602004113 pmid: 27325772 |
| [10] | Xu F, He S B, Zhang J Y, Mao Z L, Wang W X, Li T, Hua J, Du S S, Xu P B, Li L, Lian H L, Yang H Q . Photoactivated CRY1 and phyB interact directly with AUX/IAA proteins to inhibit auxin signaling in Arabidopsis. Mol Plant, 2017,11:523-541 |
| [11] | Facella P, Daddiego L, Perrotta G . CRY1a influences the diurnal transcription of photoreceptor genes in tomato plants after gibberellin treatment. Plant Signal Behav, 2012,7:1034-1036 |
| [12] | Li Y Y, Mao K, Zhao C, Zhang R F, Zhao X Y, Zhang H L, Shu H R, Zhao Y . Molecular cloning of cryptochrome 1 from apple and its functional characterization in Arabidopsis. Plant Physiol Biochem, 2013,67:169-177 |
| [13] |
Zhang Y C, Gong S F, Sang F, Yang H Q . Functional and signaling mechanism analysis of rice CRYPTOCHROME 1. 2006, Plant J, 46:971-983
doi: 10.1111/j.1365-313X.2006.02753.x pmid: 16805731 |
| [14] |
Hirose F, Shinomura T, Tanabata T, Shimada H, Takano M . Involvement of rice cryptochromes in de-etiolation responses and flowering. Plant Cell Physiol, 2006,47:915-925
doi: 10.1093/pcp/pcj064 pmid: 16760221 |
| [15] |
Platten J D, Foo E, Foucher F, Hecht V, Reid J B, Weller J L . The cryptochrome gene family in pea includes two differentially expressed CRY2 genes. Plant Mol Biol, 2005,59:683-696
doi: 10.1007/s11103-005-0828-z pmid: 16244915 |
| [16] | Imaizumi T, Kanegae T, Wada M . Cryptochrome nucleocytoplasmic distribution and gene expression are regulated by light quality in the fern Adiantum capillus-veneris. Plant Cell, 2000,12:81-96 |
| [17] | Imaizumi T, Kadota A, Hasebe M, Wada M . Cryptochrome light signals control development to suppress auxin sensitivity in the moss physcomitrella patens. Plant Cell, 2002,14:373-386 |
| [18] | Meng Y Y, Li H Y . Blue light-dependent interaction between cryptochrome 2 and CIB1 regulates transcription and leaf senescence in soybean. Plant Cell, 2013,25:4405-4420 |
| [19] | Zhang Q Z, Li H Y, Li R, Hu R B, Fan C M, Chen F L, Wang Z H, Liu X, Fu Y F, Lin C T . Association of the circadian rhythmic expression of GmCRY1a with a latitudinal cline in photoperiodic flowering of soybean. Proc Natl Acad Sci USA, 2008,105:21028-21033 |
| [20] | Chatterjee M, Sharma P, Khurana J P . Cryptochrome 1 from Brassica napus is up-regulated by blue light and controls hypocotyl/stem growth and anthocyanin accumulation . Plant Physiol, 2006,141:61-74 |
| [21] |
Liu H T, Liu B, Zhao C X, Pepper M, Lin C T . The action mechanisms of plant cryptochromes. Trends Plant Sci, 2011,16:684-691
doi: 10.1016/j.tplants.2011.09.002 pmid: 3277817 |
| [22] | Ahmad M, Cashmore A R . HY4 gene of A. thaliana encodes a protein with characteristics of a blue-light photoreceptor . Nature, 1993,366:162-166 |
| [23] |
Guo H W, Yang H Y, Mockler T C, Lin C T . Regulation of flowering time by Arabidopsis photoreceptors. Science, 1998,279:1360-1363
doi: 10.1016/j.pneurobio.2009.12.005. |
| [24] | Wu G, Spalding E P . Separate functions for nuclear and cytoplasmic cryptochrome 1 during photomorphogenesis of Arabidopsis seedlings. Proc Natl Acad Sci USA, 2007,104:18813-18818 |
| [25] |
Yu X H, Klejno J, Zhao X Y, Dror S, Maskit M, Yang H Y, Janet L, Liu X M, Javier L, Lin C T . Arabidopsis cryptochrome 2 completes its posttranslational life cycle in the nucleus. Plant Cell, 2007,19:3146-3156
doi: 10.1105/tpc.107.053017 |
| [26] | Kleine T, Lockhart P, Batschauer A . An Arabidopsis protein closely related to Synechocystis cryptochrome is targeted to organelles. Plant J, 2003,35:93-103 |
| [27] | 闫蕾, 杨宗举, 苏亮, 肖阳, 郭林, 宋梅芳, 孙蕾, 孟凡华, 白建荣, 杨建平 . 2个玉米ZmCRY1a基因的克隆及其响应光质处理的表达模式. 作物学报, 2016,42:1298-1308 |
| Yan L, Yang Z J, Su L, Xiao Y, Guo L, Song M F, Sun L, Meng F H, Bai J R, Yang J P . Molecular cloning of two maize (Zea mays) CRY1a genes and their expression patterns of in response to different light treatments. Acta Agron Sin, 2016,42:1298-1308 (in Chinese with English abstract) | |
| [28] | Rajeevan M S, Ranamukhaarachi D G, Vernon S D, Unger E R . Use of real-time quantitative PCR to validate the results of cDNA array and differential display PCR technologies. Methods, 2001,25:443-451 |
| [29] | Yang Y J, Zuo Z C, Zhao X Y, Li X, John K, Lia Y, Chen P, Liang S P, Yu X H, Liu X M, Lin C T . Blue-light-independent activity of Arabidopsis cryptochromes in the regulation of steady-state levels of protein and mRNA expression. Mol Plant, 2008,1:167-177 |
| [30] |
Wang Q, Liu Q, Wang X, Zuo Z, Oka Y, Lin C . New insights into the mechanisms of phytochrome-cryptochrome coaction. New Phytol, 2018,217:547-551
doi: 10.1111/nph.14886 pmid: 29139123 |
| [31] | de Wit M, Keuskamp D H, Bongers F J, Hornitschek P, Gommers C M M, Reinen E, Martínez-Cerón C, Fankhauser C, Pierik R . Integration of phytochrome and cryptochrome signals determines plant growth during competition for light. Curr Biol, 2016,26:3320-3326 |
| [32] |
Xu P B, Lian H L, Wang W X, Xu F, Yang H Q . Pivotal roles of the phytochrome-interacting factors in cryptochrome signaling. Mol Plant, 2016,9:496-497
doi: 10.1016/j.molp.2016.02.007 pmid: 26921621 |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [4] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [5] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [6] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [7] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [8] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [9] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [10] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [11] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [12] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [13] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [14] | 张倩, 韩本高, 张博, 盛开, 李岚涛, 王宜伦. 控失尿素减施及不同配比对夏玉米产量及氮肥效率的影响[J]. 作物学报, 2022, 48(1): 180-192. |
| [15] | 苏达, 颜晓军, 蔡远扬, 梁恬, 吴良泉, MUHAMMAD AtifMuneer, 叶德练. 磷肥对甜玉米籽粒植酸和锌有效性的影响[J]. 作物学报, 2022, 48(1): 203-214. |
|
||