欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (4): 886-895.doi: 10.3724/SP.J.1006.2022.13026
刘磊( ), 詹为民(), 丁武思, 刘通, 崔连花, 姜良良, 张艳培*(), 杨建平*()
), 詹为民(), 丁武思, 刘通, 崔连花, 姜良良, 张艳培*(), 杨建平*()
LIU Lei(), ZHAN Wei-Min(), DING Wu-Si, LIU Tong, CUI Lian-Hua, JIANG Liang-Liang, ZHANG Yan-Pei*(), YANG Jian-Ping*()
摘要:
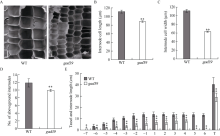
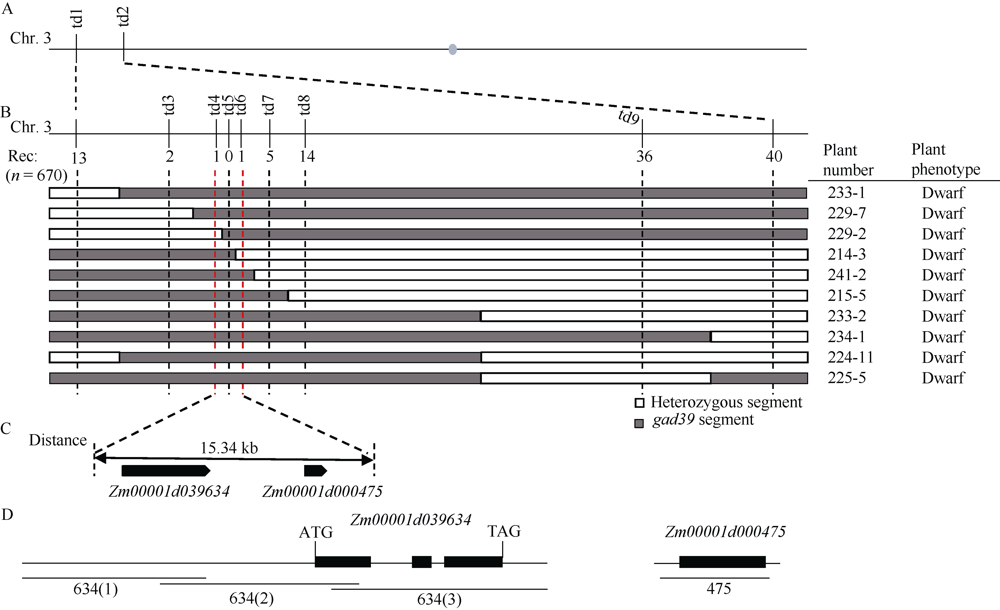
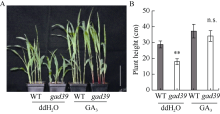
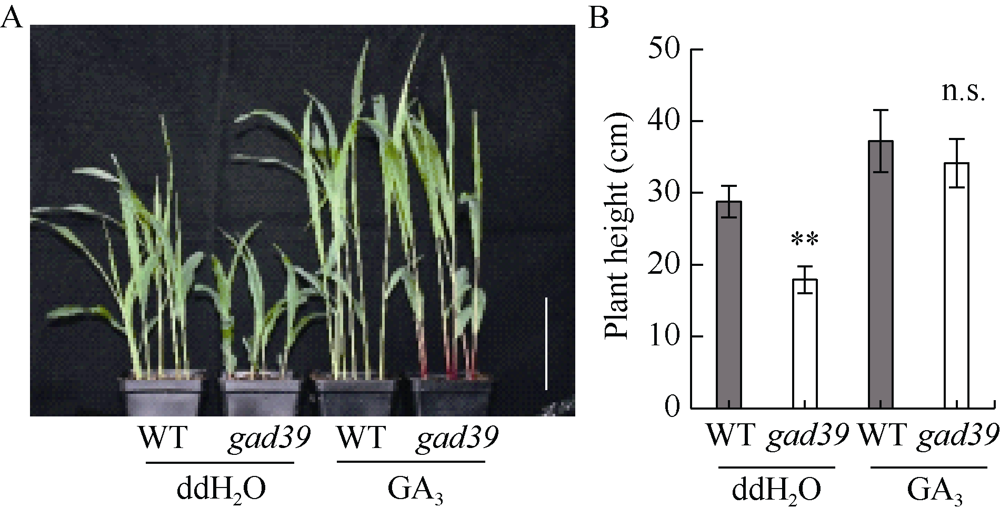
株高决定了玉米的种植密度和抗倒伏性, 进而影响产量和品质, 是玉米育种中重要的选择性状之一, 因此对控制玉米株高相关基因的遗传和分子机制的研究具有重要意义。本文对源自玉米自交系Mo17的矮化自然突变体gad39进行了表型鉴定、细胞学观察、遗传分析、基因定位和赤霉素(GA3)处理等研究。田间种植条件下, 整个生育期gad39的株高都明显矮于野生型Mo17, 吐丝期仅100.00 cm, 与野生型的192.60 cm相比, 下降了48.08%, 差异达到极显著水平; 进一步分析发现gad39的雄穗长度显著变短, 节间数目显著减少。扫描电镜观察发现, 茎秆纵向细胞的宽度和长度显著变小。雄穗变短、节间数目减少和纵向细胞变小是导致gad39植株矮化的主要原因。除植株矮化外, gad39分蘖数增加, 穗位降低, 茎秆变细, 叶片变短和雌穗变短。遗传分析表明, gad39的突变表型由1对隐性核基因控制, 将控制矮化性状的基因定位在3号染色体长臂td4和td6标记之间。这2个标记之间的物理距离为15.34 kb, 其间包含一个控制植株矮化的基因D1/ZmGA3ox2。测序发现, gad39中的D1基因具有10个InDels和21个SNPs, 导致外显子中4个氨基酸的变异。gad39的突变位点与已报道的dwarf1、d1-4、d1-6016和d1-3286突变体不同, 是D1基因一个新的等位突变体。D1/ZmGA3ox2编码GA3氧化酶(GA3-oxidase, GA3ox), 是活性GA生物合成途径中的重要酶之一。对gad39突变体施加GA3处理, 其株高恢复至野生型水平。本研究在玉米中发现一个新的矮化等位突变遗传材料, 为进一步解析玉米株高的遗传机制奠定了基础。
| [1] | Duvick D N, Smith J, Cooper M. Long-term selection in a commercial hybrid maize breeding program. Plant Breed Rev, 2004, 24:109-151. |
| [2] |
Hébert Y, Guingo E, Loudet O. The response of root/shoot partitioning and root morphology to light reduction in maize genotypes. Crop Sci, 2001, 41:363-371.
doi: 10.2135/cropsci2001.412363x |
| [3] | 何少勇. 玉米矮秆突变体的等位性鉴定及对外源激素的敏感性研究. 四川农业大学硕士学位论文,四川成都, 2017. |
| He S Y. Studies on Allelic Identification and Sensitivity to Exogenous Hormones of Maize Dwarf Mutant. MS Thesis of Sichuan Agricultural University, Chengdu, Sichuan,China, 2017 (in Chinese with English abstract). | |
| [4] |
Multani D S, Briggs S P, Chamberlin M A, Blakeslee J J, Murphy A S, Johal G S. Loss of an MDR transporter in compact stalks of maize br2 and sorghum dw3 mutants. Science, 2003, 302:81-84.
pmid: 14526073 |
| [5] |
Zhang X G, Hou X B, Liu Y H, Zheng L J, Yi Q, Zhang H J, Huang X R, Zhang J J, Hu Y F, Yu G W, Liu H M, Li Y P, Huang H H, Zhan F L, Chen L, Tang J H, Huang Y B. Maize brachytic2 (br2) suppresses the elongation of lower internodes for excessive auxin accumulation in the intercalary meristem region. BMC Plant Biol, 2019, 19:589.
doi: 10.1186/s12870-019-2200-5 |
| [6] | Hartwig T, Chuck G S, Fujioka S, Klempien A, Weizbauer R, Potluri D P, Choe S, Johal G S, Schulz B. Brassinosteroid control of sex determination in maize. Proc Natl Acad Sci USA, 2011, 108:19814-19819. |
| [7] |
Best N B, Hartwig T, Budka J, Fujioka S, Johal G, Schulz B, Dilkes B P. Nana plant2 encodes a maize ortholog of the Arabidopsis brassinosteroid biosynthesis protein Dwarf1, identifying developmental interactions between brassinosteroids and gibberellins. Plant Physiol, 2016, 171:2633-2647.
doi: 10.1104/pp.16.00399 |
| [8] |
Peng J, Richards D E, Hartley N M, Murphy G P, Devos K M, Flintham J E, Beales J, Fish J L, Worland A J, Pelica F, Sudhakar D, Christou P, Snape J W, Gale M D, Harberd N P. ‘Green revolution’ genes encode mutant gibberellin response modulators. Nature, 1999, 400:256-261.
doi: 10.1038/22307 |
| [9] |
Lawit S J, Wych H M, Xu D, Kundu S, Tomeset D T. Maize DELLA proteins dwarf plant8 and dwarf plant9 as modulators of plant development. Plant Cell Physiol, 2010, 51:1854-1868.
doi: 10.1093/pcp/pcq153 pmid: 20937610 |
| [10] | Wang Y J, Deng D X, Ding H D, Xu X M, Zhang R, Wang S X, Bian Y L, Yin Z T, Chen Y. Gibberellin biosynthetic deficiency is responsible for maize dominant Dwarf11 (D11) mutant phenotype: physiological and transcriptomic evidence. PLoS One, 2013, 8:e66466. |
| [11] | 王立静, 哈丽旦, 张素梅, 徐春花, 李启芳, 刘保申. 新的玉米矮秆突变基因的鉴定与遗传分析. 华北农学报, 2008, 23(5):23-25. |
| Wang L J, Ha L D, Zhang S M, Xu C H, Li Q F, Liu B S. Identification and genetic analysis of a novel dwarf mutant gene in maize. Acta Agric Boreali-Sin, 2008, 23(5):23-25 (in Chinese with English abstract). | |
| [12] |
Li P C, Wei J, Wang H M, Fang Y, Yin S Y, Xu Y, Liu J, Yang Z F, Xu C W. Natural variation and domestication selection of ZmPGP1 affects plant architecture and yield-related traits in maize. Genes, 2019, 10:664.
doi: 10.3390/genes10090664 |
| [13] |
Li Z X, Zhang X R, Zhao Y J, Li Y J, Zhang G F, Peng Z H, Zhang J R. Enhancing auxin accumulation in maize root tips improves root growth and dwarfs plant height. Plant Biotechnol J, 2018, 16:86-99.
doi: 10.1111/pbi.2018.16.issue-1 |
| [14] |
Li H C, Wang L J, Liu M S, Dong Z B, Li Q F, Fei S L, Xiang H T, Liu B S, Jin W W. Maize plant architecture is regulated by the ethylene biosynthetic gene ZmACS7. Plant Physiol, 2020, 183:1184-1199.
doi: 10.1104/pp.19.01421 |
| [15] |
Kir G, Ye H, Nelissen H, Neelakandan N K, Kusnandar A S, Luo A, Inzé D, Sylvester A W, Yin Y, Becraft P W. RNAi knockdown of BRI1 in maize reveals novel functions for brassinosteroid signaling in controlling plant architecture. Plant Physiol, 2015, 169:826-839.
doi: 10.1104/pp.15.00367 |
| [16] |
Castorina G, Persico M, Zilio M, Sangiorgio S, Carabelli L, Consonni G. The maize lilliputian1 (lil1) gene, encoding a brassinosteroid cytochrome P450 C-6 oxidase, is involved in plant growth and drought response. Ann Bot, 2018, 122:227-238.
doi: 10.1093/aob/mcy047 |
| [17] | Makarevitch I, Thompson A, Muehlbauer G J, Springer N M. Brd1 gene in maize encodes a brassinosteroid C-6 oxidase. PLoS One, 2012, 7:e30798. |
| [18] |
Phinney B O. Growth response of single-gene dwarf mutants in maize to gibberellic acid. Proc Natl Acad Sci USA, 1956, 42:185-189.
doi: 10.1073/pnas.42.4.185 |
| [19] |
Chen Y, Hou M M, Liu L J, Wu S, Shen Y, Ishiyama K, Kobayashi M, McCarty D R, Tan B C. The maize dwarf encodes a gibberellin 3-oxidase and is dual localized to the nucleus and cytosol. Plant Physiol, 2014, 166:2028-2039.
doi: 10.1104/pp.114.247486 pmid: 25341533 |
| [20] |
Teng F, Zhai L H, Liu R X, Bai W, Wang L Q, Huo D G, Tao Y S, Zheng Y L, Zhang Z X. ZmGA3ox2, a candidate gene for a major QTL, qPH3.1, for plant height in maize. Plant J, 2013, 73:405-416.
doi: 10.1111/tpj.12038 |
| [21] |
Winkler R G, Helentjaris T. The maize Dwarf3 gene encodes a cytochrome P450-mediated early step in gibberellin biosynthesis. Plant Cell, 1995, 7:1307-1317.
pmid: 7549486 |
| [22] |
Bensen R J, Johal G S, Crane V C, Tossberg J T, Schnable P S, Meeley R B, Briggs S P. Cloning and characterization of the maize An1 gene. Plant Cell, 1995, 7:75-84.
pmid: 7696880 |
| [23] |
Lv H K, Zheng J, Wang T Y, Fu J J, Huai J L, Min H W, Zhang X, Tian B H, Shi Y S, Wang G Y. The maize d2003, a novel allele of VP8, is required for maize internode elongation. Plant Mol Biol, 2014, 84:243-257.
doi: 10.1007/s11103-013-0129-x |
| [24] |
Avila L M, Cerrudo D, Swanton C, Lukens L. Brevis plant1, a putative inositol polyphosphate 5-phosphatase, is required for internode elongation in maize. J Exp Bot, 2016, 67:1577-1588.
doi: 10.1093/jxb/erv554 pmid: 26767748 |
| [25] |
Smith L G, Gerttula S M, Levy H J. Tangled1: a microtubule binding protein required for the spatial control of cytokinesis in maize. J Cell Biol, 2001, 152:231-236.
pmid: 11149933 |
| [26] |
Wang Y J, Zhao J, Lu W J, Deng D X. Gibberellin in plant height control: old player, new story. Plant Cell Rep, 2017, 36:391-398.
doi: 10.1007/s00299-017-2104-5 |
| [27] | 李祖亮. 玉米矮化突变体gad5表型分析和基因克隆. 河南大学硕士学位论文,河南开封, 2015. |
| Li Z L. Phenotype Analysis and Gene Cloning of Maize Dwarf Mutant gad5. MS Thesis of Henan University, Kaifeng, Henan,China, 2015 (in Chinese with English abstract). | |
| [28] | 李巧峡, 张丽, 王玉, 黄小霞. 赤霉素调控植物开花及花器官发育的研究进展. 中国细胞生物学学报, 2019, 41:746-758. |
| Li Q X, Zhang L, Wang Y, Huang X X. Research progress of gibberellin regulation of flowering and flower organ development in plants. Chin J Cell Biol, 2019, 41:746-758 (in Chinese with English abstract). | |
| [29] | 任晓松, 王子沐, 焦健, 田礼欣, 刘赵月, 李晶. GA处理对低温胁迫条件下玉米种子呼吸代谢的影响. 生态学杂志, 2020, 39:847-854. |
| Ren X S, Wang Z M, Jiao J, Tian L X, Liu Z Y, Li J. Effects of GA treatment on respiration metabolism of maize seeds under low temperature stress. Chin J Ecol, 2020, 39:847-854 (in Chinese with English abstract). | |
| [30] |
Hu S L, Wang C L, Sanchez D L, Lipk A E, Liu P, Yin Y H, Blanco M, Lübbersted T. Gibberellins promote brassinosteroids action and both increase heterosis for plant height in maize (Zea mays L.). Front Plant Sci, 2017, 8:1039.
doi: 10.3389/fpls.2017.01039 |
| [31] |
高秀华, 傅向东. 赤霉素信号转导及其调控植物生长发育的研究进展. 生物技术通报, 2018, 34(7):1-13.
doi: 10.13560/j.cnki.biotech.bull.1985.2018-0447 |
| Gao X H, Fu X D. Research progress of gibberellin signal transduction and its regulation of plant growth and development. Biotechnol Bull, 2018, 34(7):1-13 (in Chinese with English abstract). | |
| [32] |
Calderon U A, Dellaporta S L. Cell death and cell protection genes determine the fate of pistils in maize. Development, 1999, 126:435-441.
pmid: 9876173 |
| [33] | 杨睿, 张正, 杨丽莉, 张彦琴, 董春林, 常建忠. 玉米矮秆突变体A5的表型鉴定及转录组分析. 山西大学学报(自然科学版), 2020, 43:597-603. |
| Yang R, Zhang Z, Yang L L, Zhang Y Q, Dong C L, Chang J Z. Phenotype identification and transcriptome analysis of dwarf mutant A5 in maize. J Shanxi Univ (Nat Sci Edn), 2020, 43:597-603 (in Chinese with English abstract). | |
| [34] | 王武全, 曹本高, 员海燕. 玉米矮秆突变体的激素敏感性分析. 西北农林科技大学学报(自然科学版), 2017, 45(8):51-55. |
| Wang W Q, Cao B G, Yun H Y. Analysis of hormone sensitivity of maize dwarf mutant. J Northwest Agric For Univ(Nat Sci Edn), 2017, 45(8):51-55 (in Chinese with English abstract). | |
| [35] | 王关林, 方宏筠. 植物基因工程(第2版). 北京: 科学出版社, 2002. pp 742-744. |
| Wang G L, Fang H Y. Plant Gene Engineering, 2nd edn. Beijing: Science Press, 2002. pp 742-744(in Chinese). | |
| [36] |
Michelmore R W, Paran I, Kesseli R V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88:9828-9832.
doi: 10.1073/pnas.88.21.9828 |
| [37] | 徐幸. 种植密度对不同株高玉米品种茎秆抗倒伏性能及产量的影响. 吉林农业大学硕士学位论文,吉林长春, 2019. |
| Xu X. Effects of Planting Density on Stem Lodging Resistance and Yield of Different Maize Cultivars with Different Height. MS Thesis of Jilin Agricultural University, Changchun, Jilin,China, 2019 (in Chinese with English abstract). | |
| [38] | Chen Y, Tan B C. New insight in the gibberellin biosynthesis and signal transduction. Plant Signal Behav, 2015, 10:e1000140. |
| [39] |
Yamaguchi S. Gibberellin metabolism and its regulation. Annu Rev Plant Biol, 2008, 59:225-251.
doi: 10.1146/annurev.arplant.59.032607.092804 pmid: 18173378 |
| [40] |
Sun T P, Kamiya Y. The Arabidopsis GA1 locus encodes the cyclase ent-kaurene synthetase a of gibberellin biosynthesis. Plant Cell, 1994, 6:1509-1518.
pmid: 7994182 |
| [41] |
Helliwell C A, Sullivan J A, Mould R M, Gray G C, Peacock W J, Dennis E S. A plastid envelope location of Arabidopsis ent-kaurene oxidase links the plastid and endoplasmic reticulum steps of the gibberellin biosynthesis pathway. Plant J, 2001, 28:201-208.
pmid: 11722763 |
| [42] | 李强, 吴建明, 梁和, 黄杏, 丘立杭. 高等植物赤霉素生物合成及其信号转导途径. 生物技术通报, 2014, (10):16-22. |
| Li Q, Wu J M, Liang H, Huang X, Qiu L H. Synthesis and signaling pathways of gibberellin biosynthesis in higher plants. Biotechnol Bull, 2014, (10):16-22 (in Chinese with English abstract). |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 郑崇珂, 周冠华, 牛淑琳, 和亚男, 孙伟, 谢先芝. 水稻早衰突变体esl-H5的表型鉴定与基因定位[J]. 作物学报, 2022, 48(6): 1389-1400. |
| [4] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [5] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [6] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [7] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [8] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [9] | 周慧文, 丘立杭, 黄杏, 李强, 陈荣发, 范业赓, 罗含敏, 闫海锋, 翁梦苓, 周忠凤, 吴建明. 甘蔗赤霉素氧化酶基因ScGA20ox1的克隆及功能分析[J]. 作物学报, 2022, 48(4): 1017-1026. |
| [10] | 王好让, 张勇, 于春淼, 董全中, 李微微, 胡凯凤, 张明明, 薛红, 杨梦平, 宋继玲, 王磊, 杨兴勇, 邱丽娟. 大豆突变体ygl2黄绿叶基因的精细定位[J]. 作物学报, 2022, 48(4): 791-800. |
| [11] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [12] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [13] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [14] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [15] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
|
||