欢迎访问作物学报,今天是
作物学报 ›› 2018, Vol. 44 ›› Issue (9): 1301-1310.doi: 10.3724/SP.J.1006.2018.01301
龙海馨( ),邱海阳,MuhammadUZAIR,房静静,赵金凤,李学勇()
),邱海阳,MuhammadUZAIR,房静静,赵金凤,李学勇()
Hai-Xin LONG(),Hai-Yang QIU,UZAIR Muhammad,Jing-Jing FANG,Jin-Feng ZHAO,Xue-Yong LI()
摘要:
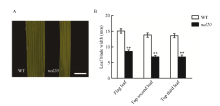
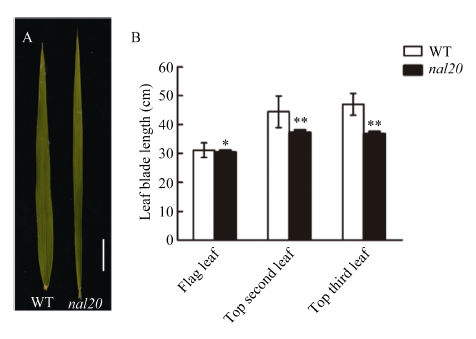
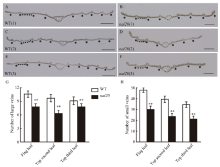
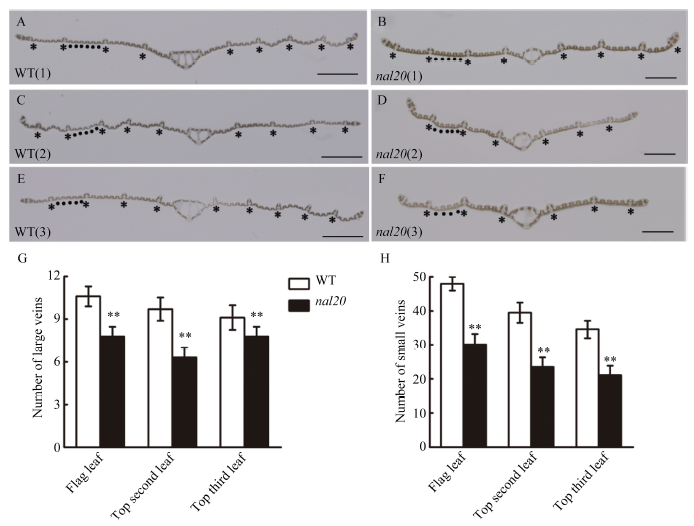
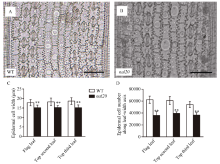
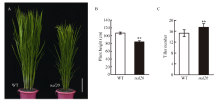
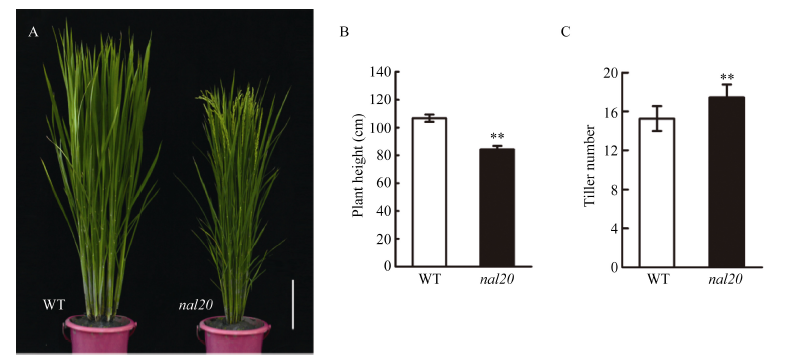
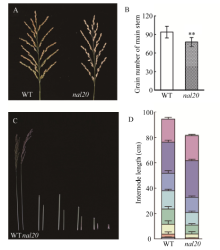
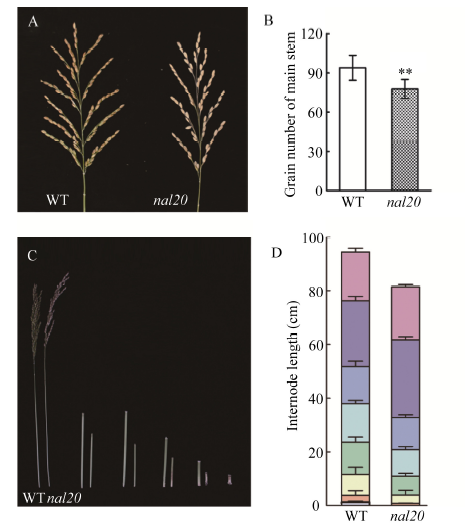
叶片作为植物光合作用的主要器官, 其面积的大小影响着光能利用率和最终产量。为了研究水稻叶片形态建成的分子机制, 利用 60Co-γ射线诱变粳稻品种春江06, 在M2代中得到1份窄叶突变体, 命名为narrow leaf20 (nal20)。该突变体叶片变窄、株高降低、分蘖增多、茎节间缩短、抽穗期提前。本研究重点调查了3片功能叶的形态, 发现突变体叶片宽度减少了50%, 叶片长度变化较小。细胞学观察表明, 叶片变窄主要是由于表皮细胞数目的减少, 而细胞大小变化不大。遗传分析表明, 该突变体表型受1对隐性基因控制。利用具有多态性的InDel分子标记及nal20与Dular配制的F2定位群体, 将该基因定位于第7染色体着丝粒区1.9 Mb范围内。二代测序结果表明, 在该范围内有455 kb的大片段缺失。本研究结果为窄叶基因NAL20的克隆和功能分析奠定了良好基础, 也为水稻株型改良提供了基因资源和育种材料。
| [1] | 朱雄涛, 汪真 . 水稻高光效生理育种初探. 福建稻麦科技, 2003,21(2):14-17 |
| Zhu X T, Wang Z . Research on high photosynthetic efficiency in the physiological breeding. Fujian Sci & Technol Rice & Wheat, 2003,21(2):14-17 (in Chinese) | |
| [2] |
Bowman J L, Eshed Y, Baum S F . Establishment of polarity in angiosperm lateral organs. Trends Genet, 2002,18:134-141
doi: 10.1016/S0168-9525(01)02601-4 pmid: 11858837 |
| [3] |
Schraer H, Hunter S J . The development of plant leaves. Plant Physiol, 2003,131:389
doi: 10.1104/pp.015347 pmid: 12586863 |
| [4] |
Murray M G, Thompson W F . Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980,8:4321-4325
doi: 10.1093/nar/8.19.4321 pmid: 324241 |
| [5] |
David-Schwartz R, Koenig D, Sinha N R . LYRATE is a key regulator of leaflet initiation and lamina outgrowth in tomato. Plant Cell, 2009,21:3093-3104
doi: 10.1105/tpc.109.069948 pmid: 2782284 |
| [6] | 陈代波, 程式华, 曹立勇 . 水稻窄叶性状的研究进展. 中国稻米, 2010,16(3):1-4 |
| Chen D B, Cheng S H, Cao L Y . Research progress of narrow leaf characters of rice. China Rice, 2010,16(3):1-4 (in Chinese) | |
| [7] | Iri A, Perera I, Ujiie K, Ishimaru K, Hirotsu N, Kashiwagi T . Partial loss-of-function of NAL1 alters canopy photosynthesis by changing the contribution of upper and lower canopy leaves in rice. Sci Rep, 2017,7(1):15958 |
| [8] | Qi J, Qian Q, Bu Q, Li S, Chen Q, Sun J, Liang W, Zhou Y, Chu C . Mutation of the rice narrow leaf1 gene, which encodes a novel protein, affects vein patterning and polar auxin transport. Plant Physiol, 2008,147:1947 |
| [9] | Woo Y M, Park H J , Su’Udi M, Yang J I, Park J J, Back K. Constitutively wilted 1, a member of the rice YUCCA gene family, is required for maintaining water homeostasis and an appropriate root to shoot ratio. Plant Mol Biol, 2007,65:125-136 |
| [10] | Fujino K, Matsuda Y, Ozawa K, Nishimura T, Koshiba T, Fraaije M W, Sekiguchi H . NARROW LEAF 7 controls leaf shape mediated by auxin in rice. Mol Genet Genomics, 2008,279:499-507 |
| [11] | Sakamoto T, Morinaka Y, Inukai Y, Kitano H, Fujioka S . Auxin signal transcription factor regulates expression of the brassinosteroid receptor gene in rice. Plant J, 2013,73:676-688 |
| [12] |
Li M, Xiong G, Li R, Cui J, Tang D, Zhang B, Ding T, Zhang B C, Pauly M, Cheng Z K, Zhou Y H . Rice cellulose synthase-like D4 is essential for normal cell-wall biosynthesis and plant growth. Plant J, 2009,60:1055-1069
doi: 10.4161/psb.5.2.10369 pmid: 19765235 |
| [13] |
Wu C, Fu Y, Hu G, Si H, Cheng S, Liu W . Isolation and characterization of a rice mutant with narrow and rolled leaves. Planta, 2010,232:313-324
doi: 10.1007/s00425-010-1180-3 pmid: 20443024 |
| [14] | Hu J, Zhu L, Zeng D, Gao Z, Guo L, Fang Y, Zhang G, Dong G, Yan M, Liu J . Identification and characterization of NARROW AND ROLLED LEAF 1, a novel gene regulating leaf morphology and plant architecture in rice. Plant Mol Biol, 2010,73:283-292 |
| [15] | Cho S H, Yoo S C, Zhang H, Pandeya D, Koh H J, Hwang J Y, Kim G T, Paek N C . The rice narrow leaf2 and narrow leaf3 loci encode WUSCHEL-related homeobox 3A (OsWOX3A) and function in leaf, spikelet, tiller and lateral root development. New Phytol, 2013,198:1071-1084 |
| [16] | Lin H, Niu L, Mchale N A, Ohme-Takagi M, Mysore K S, Tadege M . Evolutionarily conserved repressive activity of WOX proteins mediates leaf blade outgrowth and floral organ development in plants. Proc Natl Acad Sci USA, 2013,110:366-371 |
| [17] |
Ishiwata A, Ozawa M, Nagasaki H, Kato M, Noda Y, Yamaguchi T, Nosaka M, Shimizu-Sato S, Nagasaki A, Maekawa M . Two WUSCHEL-related homeobox genes, narrow leaf2 and narrow leaf3, control leaf width in rice. Plant Cell Physiol, 2013,54:779-792
doi: 10.1093/pcp/pct032 pmid: 23420902 |
| [18] | Zheng M, Wang Y, Liu X, Sun J, Wang Y, Xu Y, Xu J L, Long W, Zhu X, Guo X . The RICE MINUTE-LIKE1 (RML1) gene, encoding a ribosomal large subunit protein L3B, regulates leaf morphology and plant architecture in rice. J Exp Bot, 2016,67:3457-3469 |
| [19] | Yang W, Guo Z, Huang C, Wang K, Jiang N, Feng H, Chen G, Liu Q, Xiong L . Genome-wide association study of rice (Oryza sativa L.) leaf traits with a high-throughput leaf scorer. J Exp Bot, 2015,66:5605-5615 |
| [20] | Yoshikawa T, Eiguchi M, Hibara K I, Ito J I, Nagato Y . Rice SLENDER LEAF 1 gene encodes cellulose synthase-like D4 and is specifically expressed in M-phase cells to regulate cell proliferation. J Exp Bot, 2013,64:2049 |
| [21] | Jiang D, Fang J, Lou L, Zhao J, Yuan S, Yin L, Sun W, Peng L, Guo B, Li X . Characterization of a null allelic mutant of the rice NAL1 gene reveals its role in regulating cell division. PLoS One, 2015,10:e0118169 |
| [22] | Michelmore R W, Paran I, Kesseli R V . Identification of markers linked to disease-resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991,88:9828 |
| [23] |
Xue W, Xing Y, Weng X, Zhao Y, Tang W, Wang L, Zhou H, Yu S, Xu C, Li X . Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat Genet, 2009,40:761-767
doi: 10.1038/ng.143 pmid: 18454147 |
| [24] | 谈聪, 翁小煜, 鄢文豪, 白旭峰, 邢永忠 . 多效性基因Ghd7调控水稻剑叶面积. 遗传, 2012,34:901-906 |
| Tan C, Weng X Y, Yan W H, Bai X F, Xing Y Z . Ghd7, a pleiotropic gene controlling flag leaf erea in rice. Hereditas(Beijing), 2012,34:901-906 (in Chinese with English abstract) | |
| [25] | Li L, Yan W, Xue W, Shao D, Xing Y . Evolution and association analysis of Ghd7 in rice. PLoS One, 2012,7:e34021 |
| [26] | Doi K, Izawa T, Fuse T, Yamanouchi U, Kubo T, Shimatani Z, Yano M, Yoshimura A . Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1 . Genes Dev, 2004,18:926-936 |
| [27] |
Hayama R, Yokoi S, Tamaki S, Yano M, Shimamoto K . Adaptation of photoperiodic control pathways produces short-day flowering in rice. Nature, 2003,422:719-722
doi: 10.1038/nature01549 pmid: 12700762 |
| [28] | 肖珂, 左海龙, 巩迎军, 张俊芝, 张永娟, 董彦君 . 控制水稻剑叶形态相关性状的数量基因位点(QTL)的定位. 上海师范大学学报(自然科学版), 2007,36(2):66-70 |
| Xiao K, Zuo H L, Gong Y J, Zhang J Z, Zhang Y J, Dong Y J . Locating quantitative trait loci affecting flag-leaf shape traits in rice (Oryza sativa L.).J Shanghai Norm Univ(Nat Sci), 2007,36(2):66-70 (in Chinese with English abstract) |
| [1] | 田甜, 陈丽娟, 何华勤. 基于Meta-QTL和RNA-seq的整合分析挖掘水稻抗稻瘟病候选基因[J]. 作物学报, 2022, 48(6): 1372-1388. |
| [2] | 郑崇珂, 周冠华, 牛淑琳, 和亚男, 孙伟, 谢先芝. 水稻早衰突变体esl-H5的表型鉴定与基因定位[J]. 作物学报, 2022, 48(6): 1389-1400. |
| [3] | 周文期, 强晓霞, 王森, 江静雯, 卫万荣. 水稻OsLPL2/PIR基因抗旱耐盐机制研究[J]. 作物学报, 2022, 48(6): 1401-1415. |
| [4] | 郑小龙, 周菁清, 白杨, 邵雅芳, 章林平, 胡培松, 魏祥进. 粳稻不同穗部籽粒的淀粉与垩白品质差异及分子机制[J]. 作物学报, 2022, 48(6): 1425-1436. |
| [5] | 颜佳倩, 顾逸彪, 薛张逸, 周天阳, 葛芊芊, 张耗, 刘立军, 王志琴, 顾骏飞, 杨建昌, 周振玲, 徐大勇. 耐盐性不同水稻品种对盐胁迫的响应差异及其机制[J]. 作物学报, 2022, 48(6): 1463-1475. |
| [6] | 杨建昌, 李超卿, 江贻. 稻米氨基酸含量和组分及其调控[J]. 作物学报, 2022, 48(5): 1037-1050. |
| [7] | 杨德卫, 王勋, 郑星星, 项信权, 崔海涛, 李生平, 唐定中. OsSAMS1在水稻稻瘟病抗性中的功能研究[J]. 作物学报, 2022, 48(5): 1119-1128. |
| [8] | 朱峥, 王田幸子, 陈悦, 刘玉晴, 燕高伟, 徐珊, 马金姣, 窦世娟, 李莉云, 刘国振. 水稻转录因子WRKY68在Xa21介导的抗白叶枯病反应中发挥正调控作用[J]. 作物学报, 2022, 48(5): 1129-1140. |
| [9] | 王小雷, 李炜星, 欧阳林娟, 徐杰, 陈小荣, 边建民, 胡丽芳, 彭小松, 贺晓鹏, 傅军如, 周大虎, 贺浩华, 孙晓棠, 朱昌兰. 基于染色体片段置换系群体检测水稻株型性状QTL[J]. 作物学报, 2022, 48(5): 1141-1151. |
| [10] | 王泽, 周钦阳, 刘聪, 穆悦, 郭威, 丁艳锋, 二宫正士. 基于无人机和地面图像的田间水稻冠层参数估测与评价[J]. 作物学报, 2022, 48(5): 1248-1261. |
| [11] | 陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展[J]. 作物学报, 2022, 48(4): 781-790. |
| [12] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [13] | 王吕, 崔月贞, 吴玉红, 郝兴顺, 张春辉, 王俊义, 刘怡欣, 李小刚, 秦宇航. 绿肥稻秆协同还田下氮肥减量的增产和培肥短期效应[J]. 作物学报, 2022, 48(4): 952-961. |
| [14] | 巫燕飞, 胡琴, 周棋, 杜雪竹, 盛锋. 水稻延伸因子复合体家族基因鉴定及非生物胁迫诱导表达模式分析[J]. 作物学报, 2022, 48(3): 644-655. |
| [15] | 陈云, 李思宇, 朱安, 刘昆, 张亚军, 张耗, 顾骏飞, 张伟杨, 刘立军, 杨建昌. 播种量和穗肥施氮量对优质食味直播水稻产量和品质的影响[J]. 作物学报, 2022, 48(3): 656-666. |
|
||