欢迎访问作物学报,今天是
作物学报 ›› 2019, Vol. 45 ›› Issue (2): 235-247.doi: 10.3724/SP.J.1006.2019.84063
邹雪1,2,丁凡1,余金龙1,彭洁2,邓孟胜2,王宇2,刘丽芳1,余韩开宗1,陈年伟1,王西瑶2,*( )
)
Xue ZOU1,2,Fan DING1,Jin-Long YU1,Jie PENG2,Meng-Sheng DENG2,Yu WANG2,Li-Fang LIU1,Kai-Zong YU-HAN1,Nian-Wei CHEN1,Xi-Yao WANG2,*()
摘要:
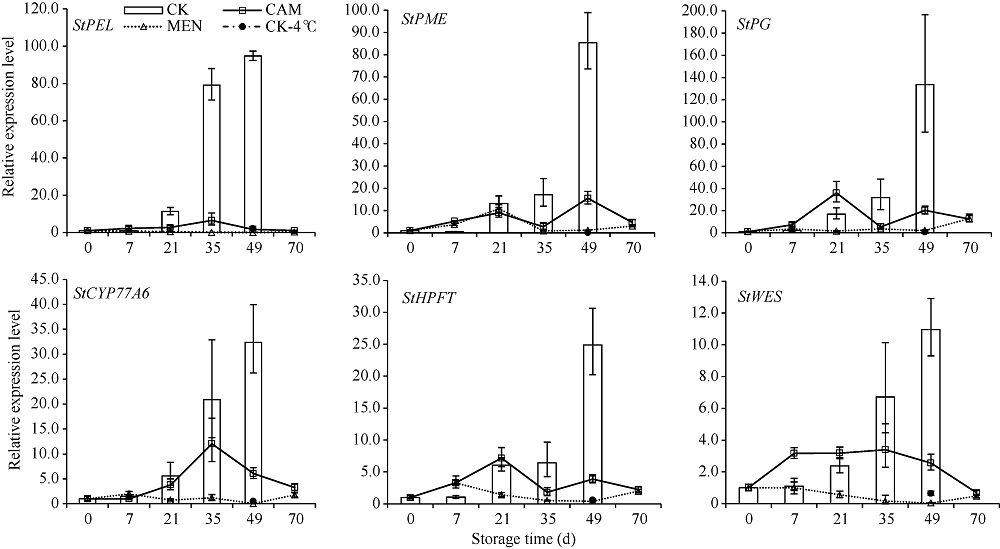
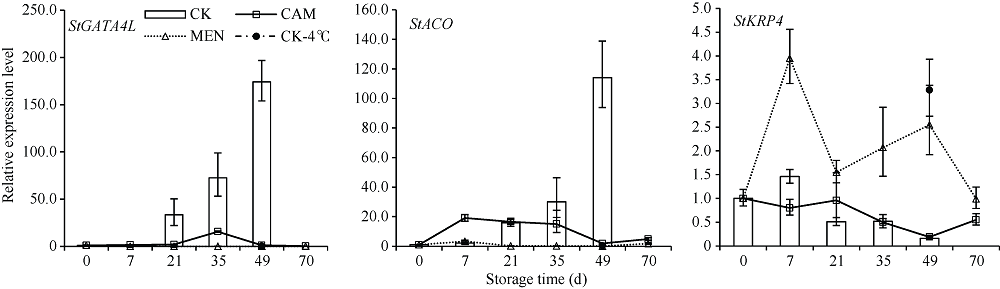
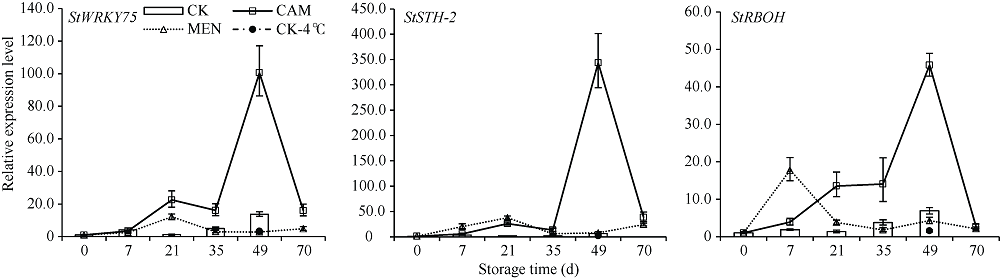
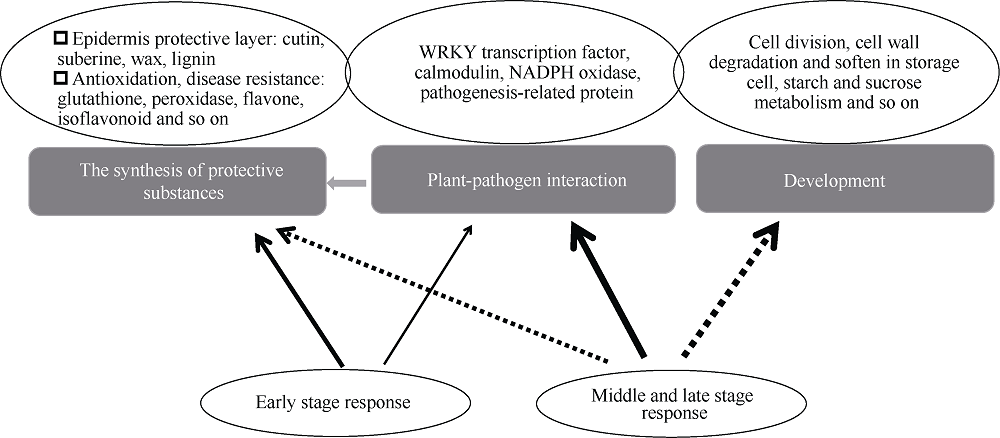
马铃薯块茎过早萌芽会降低商品价值, 本试验比较挥发性物质的抑芽效应, 并从转录和蛋白水平分析其作用机制。结果表明, 抑芽能力薄荷醇?樟脑?萘, 萌芽受抑降低代谢消耗, 薄荷醇处理180 d时的重量损失只有对照损失的36%。樟脑处理萌芽块茎3 d引起表达显著上调(或下调)的基因和蛋白分别有1227 (299)和296 (204)个, 主要参与响应刺激、防御反应。贮藏期间, 果胶代谢基因PEL、PME、PG, 角质合成基因CYP77A6、HPFT、WES, 乙烯合成基因ACO以及转录因子编码基因GATA4L的表达量随时间升高。樟脑早期不同程度地刺激这些基因表达, 中后期则抑制, 49 d时只有对照的0.68%~23.35%。薄荷醇使上述基因表达保持低水平, 但可提高细胞周期负控基因KRP4的表达, 为对照的15.9倍。植物病原菌互作通路中的基因WRKY75、STH-2、RBOH表达受樟脑诱导并在后期高表达。樟脑和薄荷醇均能抑制生长发育基因的表达, 造成芽死亡, 降低贮藏损耗。前者早期能促进合成保护性物质, 萌芽时有更强烈的抗菌反应; 后者则阻碍细胞分裂。
| [1] | 肖关丽, 郭华春 . 不同生理年龄马铃薯种薯芽中的内源激素含量变化及其对马铃薯植株生长发育的影响. 植物生理学报, 2007,43:818-820. |
| Xiao G L, Guo H C . Changes in endogenous hormone contents in bud of seed potato (Solanum tuberosum L.) with different physiological ages and its effect on growth and development. Plant Physiol J, 2007,43:818-820 (in Chinese with English abstract). | |
| [2] |
Daniels-Lake B J, Pruski K, Prange R K . Using ethylene gas and chlorpropham potato sprout inhibitors together. Potato Res, 2011,54:223-236.
doi: 10.1007/s11540-011-9188-z |
| [3] |
Vijay P, Ezekiel R, Pandey R . Use of CIPC as a potato sprout suppressant: health and environmental concerns and future options. Qual Assur Safety Crops Foods, 2018,10:17-24.
doi: 10.3920/QAS2017.1088 |
| [4] |
Gómez-Castillo D . Effects of essential oils on sprout suppression and quality of potato cultivars. Postharvest Biol Technol, 2013,82:15-21.
doi: 10.1016/j.postharvbio.2013.02.017 |
| [5] |
Hartmans K J . The use of carvone in agriculture: sprout suppression of potatoes and antifungal activity against potato tuber and other plant diseases. Ind Crops Prod, 1995,4:3-13.
doi: 10.1016/0926-6690(95)00005-W |
| [6] |
Oosterhaven K, Poolman B, Smid E J . S-carvone as a natural potato sprout inhibiting, fungistatic and bacteristatic compound. Ind Crops Prod, 1995,4:23-31.
doi: 10.1016/0926-6690(95)00007-Y |
| [7] |
Teper-Bamnolker P, Dudai N, Fischer R, Belausov E, Zemach H, Shoseyov O, Eshel D . Mint essential oil can induce or inhibit potato sprouing by differential alteration of apical meristem. Planta, 2010,232:179-186.
doi: 10.1007/s00425-010-1154-5 pmid: 20390295 |
| [8] |
Weerd J W, Thornton M K, Shafii B . Sprout suppressing residue levels of 1,4-dimethylnaphthalene (1,4DMN) in potato cultivars. Am J Potato Res, 2010,87:434-445.
doi: 10.1007/s12230-010-9146-3 |
| [9] |
Campbell M A, Gleichsner A, Alsbury R, Horvath D, Suttle J . The sprout inhibitors chlorpropham and 1,4-dimethylnaphthalene elicit different transcriptional profiles and do not suppress growth through a prolongation of the dormant state. Plant Mol Biol, 2010,73:181-189.
doi: 10.1007/s11103-010-9607-6 pmid: 20135197 |
| [10] |
Campbell M A, Gleichsner A, Hilldorfer L, Horvath D, Suttle J . The sprout inhibitor 1,4-dimethylnaphthalene induces the expression of the cell cycle inhibitors KRP1 and KRP2 in potatoes. Funct Integr Genomics, 2012,12:533-541.
doi: 10.1007/s10142-011-0257-9 pmid: 22113341 |
| [11] |
Li L Q, Zou X, Deng M S, Peng J, Huang X L, Lu X, Fang C C, Wang X Y . Comparative morphology, transcription, and proteomics study revealing the key molecular mechanism of camphor on the potato tuber sprouting effect. Int J Mol Sci, 2017,18:2280.
doi: 10.3390/ijms18112280 pmid: 29084178 |
| [12] |
Liu B L, Zhang N, Wen Y K, Jin X, Yang J W, Si H J, Wang D . Transcriptomic changes during tuber dormancy release process revealed by RNA sequencing in potato. J Biotechnol, 2015,198:17-30.
doi: 10.1016/j.jbiotec.2015.01.019 pmid: 25661840 |
| [13] |
Yang Y, Qiang X, Owsiany K, Zhang S, Thannhauser T W, Li L . Evaluation of different multidimensional LC-MS/MS pipelines for isobaric tags for relative and absolute quantitation (iTRAQ)- based proteomic analysis of potato tubers in response to cold storage. J Proteome Res, 2011,10:4647-4660.
doi: 10.1021/pr200455s pmid: 21842911 |
| [14] |
The Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature, 2011,475:189-195.
doi: 10.1038/nature10158 pmid: 21743474 |
| [15] |
Mortazavi A, Williams B A, McCue K, Schaeffer L, Wold B . Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat Methods, 2008,5:621-628.
doi: 10.1038/nmeth.1226 |
| [16] |
邹雪, 邓孟胜, 李立芹, 余金龙, 丁凡, 黄雪丽, 彭洁, 帅禹, 蔡诚诚, 王西瑶 . 油菜素内酯合成和信号转导基因在马铃薯块茎贮藏期间的表达变化及对萌芽的影响. 作物学报, 2017,43:811-820.
doi: 10.3724/SP.J.1006.2017.00811 |
|
Zou X, Deng M S, Li L Q, Yu J L, Ding F, Huang X L, Peng J, Shuai Y, Cai C C, Wang X Y . Expression changes of genes related to brassinosteroid biosynthesis and signal transduction during potato storage and its effect on tuber sprouting. Acta Agron Sin, 2017,43:811-820 (in Chinese with English abstract).
doi: 10.3724/SP.J.1006.2017.00811 |
|
| [17] |
Dixon R A . Natural products and plant disease resistance. Nature, 2001,411:843-847.
doi: 10.1038/35081178 |
| [18] |
Kobayashi M, Ohura I, Kawakita K, Yokota N, Fujiwara M, Shimamoto K, Doke N, Yoshioka H . Calcium-dependent protein kinases regulate the production of reactive oxygen species by potato NADPH oxidase. Plant Cell, 2007,19:1065-1080.
doi: 10.1105/tpc.106.048884 pmid: 17400895 |
| [19] |
Baydar H, Karadoğan T . The effects of volatile oils on in vitro potato sprout growth. Potato Res, 2003,46:1-8.
doi: 10.1007/BF02736098 |
| [20] |
马进, 郑钢, 裴翠明, 张振亚 . 基于iTRAQ质谱分析技术筛选南方型紫花苜蓿根部响应盐胁迫差异表达蛋白. 农业生物技术学报, 2016,24:497-509.
doi: 10.3969/j.issn.1674-7968.2016.04.004 |
|
Ma J, Zheng G, Pei C M, Zhang Z Y . Screening differentially expressed proteins in southern type alfalfa (Medicago sativa ‘Millenium’) root upon salt stress by iTRAQ protein mass spectrometry. J Agric Biotechnol, 2016,24:497-509 (in Chinese with English abstract).
doi: 10.3969/j.issn.1674-7968.2016.04.004 |
|
| [21] | Takahashi K, Niwa H, Yokota N, Kubota K, Inoue H . Widespread tissue expression of nepenthesis-like aspartic protease genes in Arabidopsis thaliana. Plant Physiol Biol, 2008,46:724-729. |
| [22] | 李合生 . 现代植物生理学(第3版). 北京: 高等教育出版社, 2012. pp 245-246. |
| Li H S . Modern Plant Physiology, 3rd edn. Beijing: Higher Education Press, 2012. pp 245-246(in Chinese). | |
| [23] |
Coleman W K, Dioxide C . Oxygen and ethylene effects on potato tuber dormancy release and sprout growth. Ann Bot, 1998,82:21-27
doi: 10.1006/anbo.1998.0645 |
| [24] |
Hartmann A, Senning M, Hedden P, Sonnewald U, Sonnewald S . Reactivation of meristem activity and sprout growth in potato tubers require both cytokinin and gibberellin. Plant Physiol, 2011,155:776-796.
doi: 10.1104/pp.110.168252 pmid: 21163959 |
| [25] |
Külen O, Stushnoff C, Davidson R D, Holm D G . Gibberelllic acid and ethephon alter potato minituber bud dormancy and improve seed tuber yield. Am J Potato Res, 2011,88:167-174.
doi: 10.1007/s12230-010-9178-8 |
| [26] |
Amita C, Sane V A, Nath P . Differential expression of pectate lyase during ethylene-induced postharvest softening of mango (Mangifera indica var. Dashehari). Physiol Planta, 2006,128:546-555.
doi: 10.1111/j.1399-3054.2006.00752.x |
| [27] |
Singh A P, Pandey S P, Rajluxmi, Pandey S, Nath P, Sane A P . Transcriptional activation of a pectate lyase gene,RbPel1, during petal abscission in rose. Postharvest Biol Technol, 2011,60:143-148.
doi: 10.1016/j.postharvbio.2010.12.014 |
| [28] |
Ranftl Q L, Bastakis E, Klermund C, Schwechheimer C . LLM- domain containing B-GATA factors control different aspects of cytokinin-regulated development in Arabidopsis thaliana. Plant Physiol, 2016,170:2295-2311.
doi: 10.1104/pp.15.01556 pmid: 26829982 |
| [29] |
Luo X M, Lin W H, Zhu S W, Zhu J Y, Sun Y, Fan X Y, Cheng M L, Hao Y Q, Oh E, Tian M M, Liu L J, Zhang M, Xie Q, Chong K, Wang Z Y . Integration of light- and brassinosteroid-signaling pathways by a GATA transcription factor in Arabidopsis. Dev Cell, 2010,19:872-883.
doi: 10.1016/j.devcel.2010.10.023 pmid: 3022420 |
| [30] |
Shin J M, Chung K M, Sakamoto S, Kojima S, Yeh C M, Ikeda M, Mitsuda N, Ohme-Takagi M . The chimeric repressor for the GATA4 transcription factor improves tolerance to nitrogen deficiency in Arabidopsis. Plant Biotechnol, 2017,34:151-158.
doi: 10.5511/plantbiotechnology.17.0727a |
| [31] | Leene J V, Hollunder J, Eeckhout D, Persiau G, Slijke E V D, Stals H . Targeted interactomics reveals a complex core cell cycle machinery in Arabidopsis thaliana. Mol Sys Biol, 2010,6:397. |
| [32] |
Sang E J, Yoko O, Jaesung N, Masaaki U, Gyung-Tae K . Kip-related protein 3 is required for control of endoreduplication in the shoot apical meristem and leaves of Arabidopsis. Mol Cells, 2013,35:47-53.
doi: 10.1007/s10059-013-2270-4 pmid: 23314608 |
| [33] |
Cheng Y, Cao L, Wang S, Li Y P, Shi X Z . Downregulation of multiple CDK inhibitor ICK/KRP genes upregulates the E2F pathway and increases cell proliferation, and organ and seed sizes in Arabidopsis. Plant J, 2013,75:642-655.
doi: 10.1111/tpj.12228 pmid: 23647236 |
| [34] |
Constabel C P, Brisson N . The defense-related STH-2 gene product of potato shows race-specific accumulation after inoculation with low concentrations of Phytophthora infestans zoospores. Planta, 1992,188:289-295.
doi: 10.1007/BF00192794 pmid: 24178317 |
| [35] |
Constabel C P, Bertrand C, Brisson N . Transgenic potato plants overexpressing the pathogenesis-related STH-2 gene show unaltered susceptibility to Phytophthora infestans and potato virus X. Plant Mol Biol, 1993,22:775-782.
doi: 10.1007/BF00027364 pmid: 8358029 |
| [36] |
El-Banna A, Hajirezaei M R, Wissing J, Ali Z, Vaas L, Heine-Dobbernack E, Jacobsen H J, Schumacher H M, Kiesecker H . Over-expression of PR-10a leads to increased salt and osmotic tolerance in potato cell cultures. J Biotechnol, 2010,150:277-287.
doi: 10.1016/j.jbiotec.2010.09.934 pmid: 20854851 |
| [37] | Park S, Gupta R, Krishna R, Kim S T, Lee D Y, Hwang D J, Bae S C, Ahn I P . Proteome analysis of disease resistance against Ralstonia solanacearum in potato cultivar CT206-10. Plant Pathol J, 2016,32:25-32. |
| [1] | 王海波, 应静文, 何礼, 叶文宣, 涂卫, 蔡兴奎, 宋波涛, 柳俊. rDNA和端粒重复序列鉴定马铃薯和茄子体细胞杂种染色体丢失和融合[J]. 作物学报, 2022, 48(5): 1273-1278. |
| [2] | 石艳艳, 马志花, 吴春花, 周永瑾, 李荣. 垄作沟覆地膜对旱地马铃薯光合特性及产量形成的影响[J]. 作物学报, 2022, 48(5): 1288-1297. |
| [3] | 冯亚, 朱熙, 罗红玉, 李世贵, 张宁, 司怀军. 马铃薯StMAPK4响应低温胁迫的功能解析[J]. 作物学报, 2022, 48(4): 896-907. |
| [4] | 张霞, 于卓, 金兴红, 于肖夏, 李景伟, 李佳奇. 马铃薯SSR引物的开发、特征分析及在彩色马铃薯材料中的扩增研究[J]. 作物学报, 2022, 48(4): 920-929. |
| [5] | 谭雪莲, 郭天文, 胡新元, 张平良, 曾骏, 刘晓伟. 黄土高原旱作区马铃薯连作根际土壤微生物群落变化特征[J]. 作物学报, 2022, 48(3): 682-694. |
| [6] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [7] | 余慧芳, 张卫娜, 康益晨, 范艳玲, 杨昕宇, 石铭福, 张茹艳, 张俊莲, 秦舒浩. 马铃薯CrRLK1Ls基因家族的鉴定及响应晚疫病菌信号的表达分析[J]. 作物学报, 2022, 48(1): 249-258. |
| [8] | 荐红举, 尚丽娜, 金中辉, 丁艺, 李燕, 王季春, 胡柏耿, Vadim Khassanov, 吕典秋. 马铃薯PIF家族成员鉴定及其对高温胁迫的响应分析[J]. 作物学报, 2022, 48(1): 86-98. |
| [9] | 许德蓉, 孙超, 毕真真, 秦天元, 王一好, 李成举, 范又方, 刘寅笃, 张俊莲, 白江平. 马铃薯StDRO1基因的多态性鉴定及其与根系性状的关联分析[J]. 作物学报, 2022, 48(1): 76-85. |
| [10] | 李玲红, 张哲, 陈永明, 尤明山, 倪中福, 邢界文. 普通小麦颖壳蜡质缺失突变体glossy1的转录组分析[J]. 作物学报, 2022, 48(1): 48-62. |
| [11] | 汪颖, 高芳, 刘兆新, 赵继浩, 赖华江, 潘小怡, 毕晨, 李向东, 杨东清. 利用WGCNA鉴定花生主茎生长基因共表达模块[J]. 作物学报, 2021, 47(9): 1639-1653. |
| [12] | 曹亮, 杜昕, 于高波, 金喜军, 张明聪, 任春元, 王孟雪, 张玉先. 外源褪黑素对干旱胁迫下绥农26大豆鼓粒期叶片碳氮代谢调控的途径分析[J]. 作物学报, 2021, 47(9): 1779-1790. |
| [13] | 黄文功, 姜卫东, 姚玉波, 宋喜霞, 刘岩, 陈思, 赵东升, 吴广文, 袁红梅, 任传英, 孙中义, 吴建忠, 康庆华. 亚麻响应低钾胁迫转录谱分析[J]. 作物学报, 2021, 47(6): 1070-1081. |
| [14] | 马贵芳, 满夏夏, 张益娟, 高豪, 孙朝霞, 李红英, 韩渊怀, 侯思宇. 谷子穗发育期转录组与叶酸代谢谱联合分析[J]. 作物学报, 2021, 47(5): 837-846. |
| [15] | 唐锐敏, 贾小云, 朱文娇, 印敬明, 杨清. 马铃薯热激转录因子HsfA3基因的克隆及其耐热性功能分析[J]. 作物学报, 2021, 47(4): 672-683. |
|
||