欢迎访问作物学报,今天是
作物学报 ›› 2018, Vol. 44 ›› Issue (02): 185-196.doi: 10.3724/SP.J.1006.2018.00185
朱莹1,**, 褚姗姗2,**, 张培培1, 程浩1, 喻德跃1, 王娇1,*
Ying ZHU1,**, Shan-Shan CHU2,**, Pei-Pei ZHANG1, Hao CHENG1, De-Yue YU1, Jiao WANG1,*
摘要:
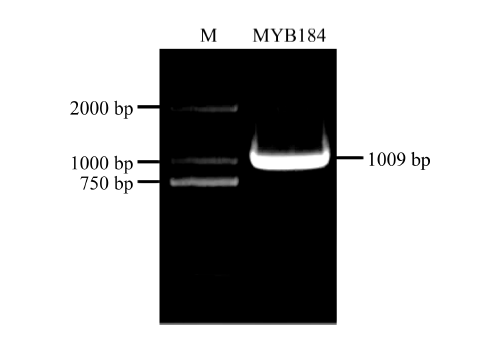
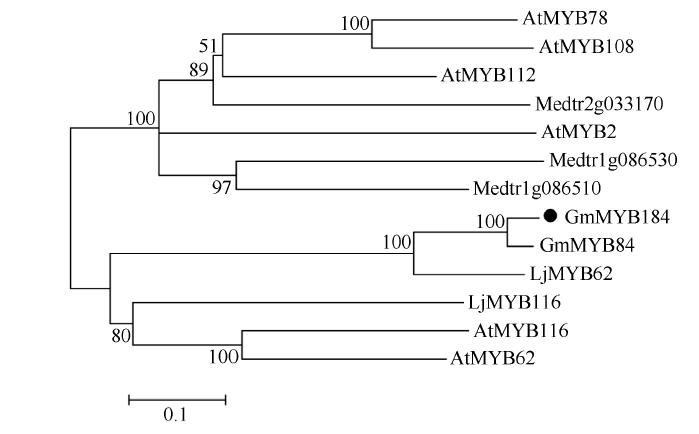
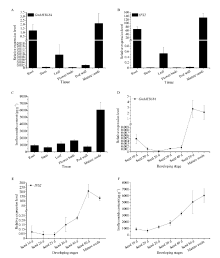
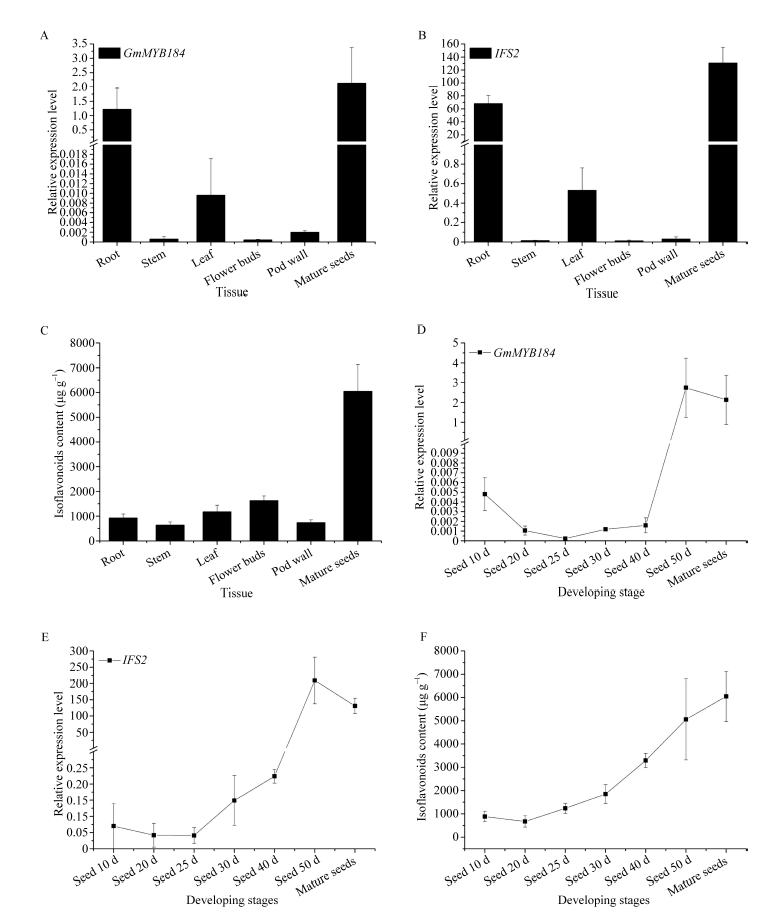
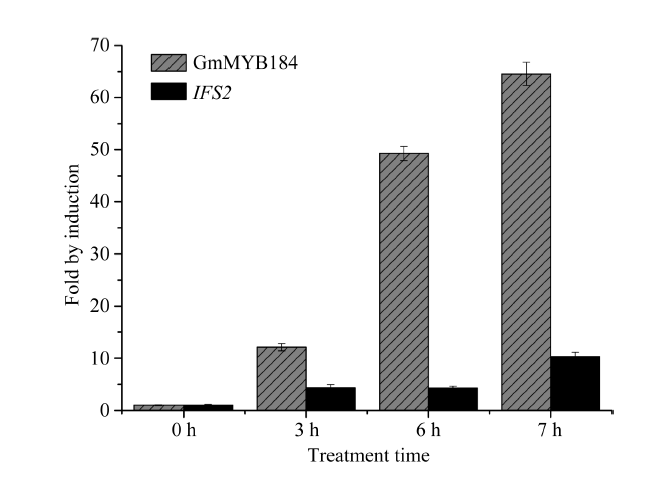
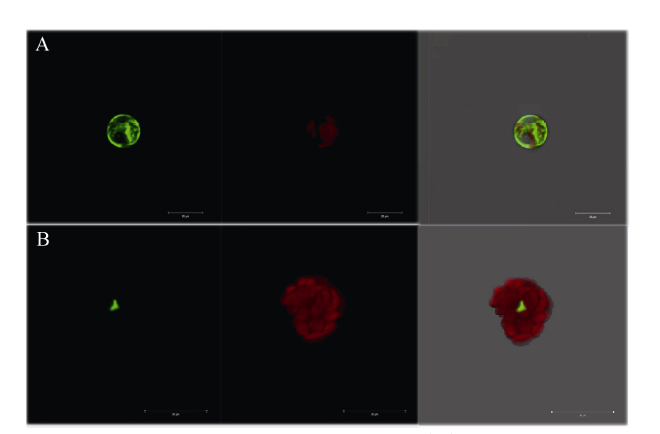
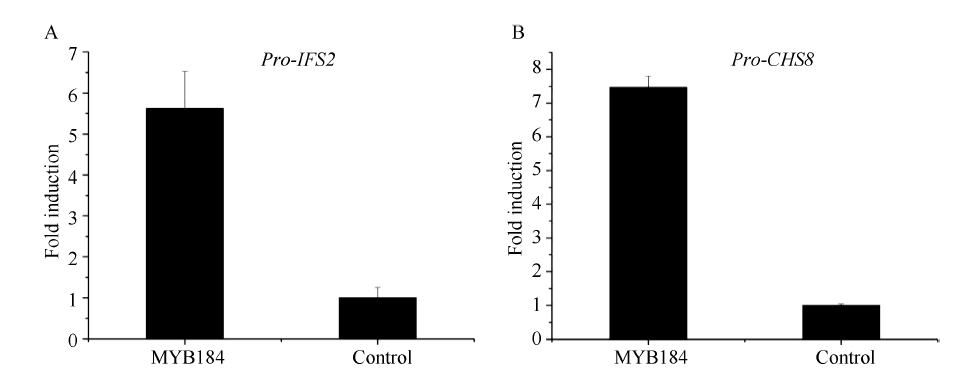
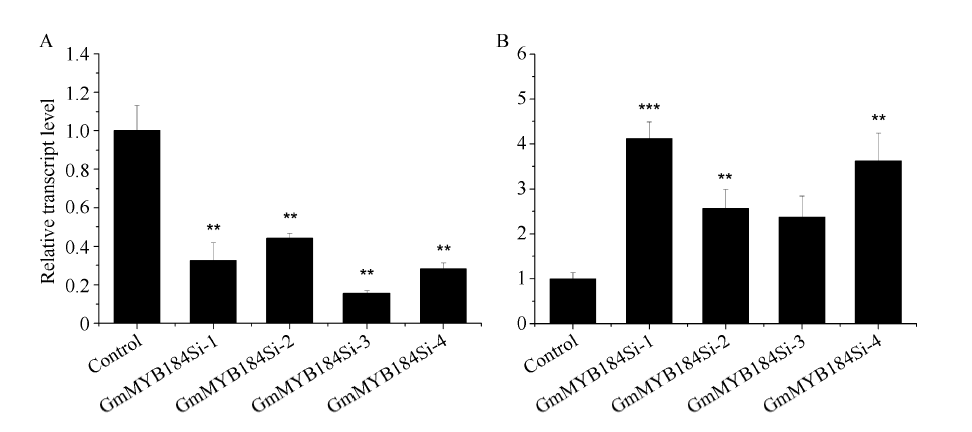
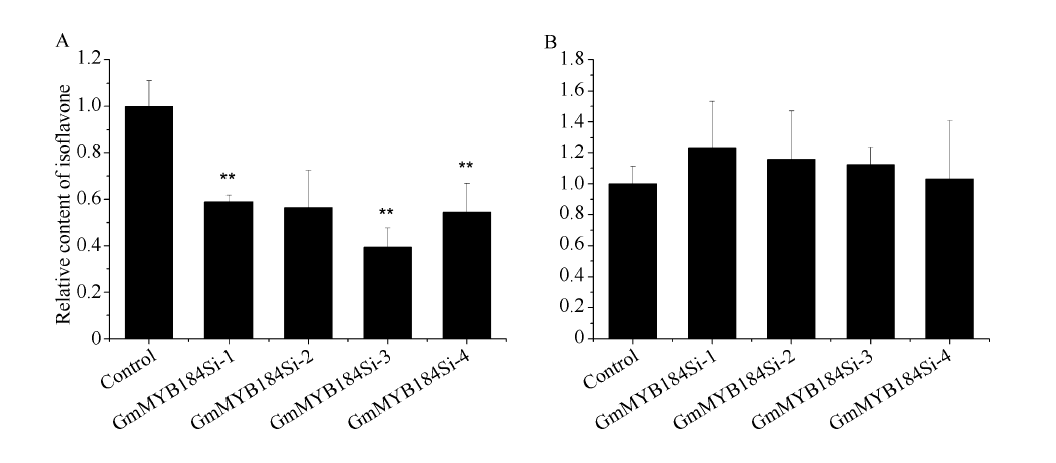
异黄酮是一类主要含在豆科植物中的次生代谢物, 在植物防御体系中发挥重要作用, 并与人类健康密切相关。大豆异黄酮含量受多基因和复杂代谢网络控制, 调控代谢途径上的结构基因不能显著改变大豆异黄酮含量, 与异黄酮代谢途径相关的转录因子的鉴定和应用可能会有效解决这个问题。本研究克隆了一个与大豆异黄酮合成相关的R2R3类型MYB转录因子GmMYB184, 并进行了初步的功能验证。亚细胞定位研究结果表明GmMYB184转录因子定位于细胞核。组织表达分析结果表明该转录因子基因与IFS2 (异黄酮合酶2编码基因)的表达模式相同。同时, GmMYB184和IFS2的表达模式与异黄酮的积累模式相似。谷胱甘肽诱导表达分析表明该转录因子基因与IFS2共同被诱导, 说明这两个基因可能参与同一或相似的生物过程。采用双荧光素酶报告系统分析其对异黄酮合成途径关键基因的转录激活活性影响, 发现GmMYB184能够促进IFS2和CHS8启动子表达活性分别提高5倍和7倍。最后, 通过发根农杆菌介导的遗传转化系统, 找到该转录因子在异黄酮合成调控中的直接作用证据。沉默GmMYB184导致大豆毛状根异黄酮含量的显著下降。但是, 过表达GmMYB184不足以显著提高毛状根中异黄酮的含量。总之, 本研究为大豆异黄酮合成分子机制探索及大豆异黄酮品质改良提供了理论依据。
| [1] | Yu O, McGonigle B. Metabolic engineering of isoflavone biosynthesis.Adv Agron 2005, 86: 147-190 |
| [2] | Cederroth C R, Nef S.Soy, phytoestrogens and metabolism: a review.Mol Cell Endocrinol 2009, 304: 30-42 |
| [3] | Rochfort S, Panozzo J.Phytochemicals for health, the role of pulses. J Agric Food Chem 2007, 55: 7981-7994 |
| [4] | Dixon R A.Natural products and plant disease resistance.Nature 2001, 411: 843-847 |
| [5] | Graham T L, Graham M Y.Signaling in soybean phenylpropanoid responses (dissection of primary, secondary, and conditioning effects of light, wounding, and elicitor treatments). Plant Physiol, 1996, 110: 1123-1133 |
| [6] | Hammerschmidt R.PHYTOALEXINS: what have we learned after 60 years? Annu Rev Phytopathol, 1999, 37: 285-306 |
| [7] | Gutierrez-Gonzalez J J, Wu X, Gillman J D, Lee J D, Zhong R, Yu O, Shannon G, Ellersieck M, Nguyen H T, Sleper D A. Intricate environment-modulated genetic networks control isoflavone accumulation in soybean seeds.BMC Plant Biol, 2010, 10: 105 |
| [8] | Dixon R A, Steele C L.Flavonoids and isoflavonoids: a gold mine for metabolic engineering.Trends Plant Sci, 1999, 4: 394-400 |
| [9] | Zernova O V, Lygin A V, Widholm J M, Lozovaya V V.Modification of isoflavones in soybean seeds via expression of multiple phenolic biosynthetic genes.Plant Physiol Biochem, 2009, 47: 769-777 |
| [10] | 易金鑫, 徐照龙, 王峻峰, 张大勇, 何晓兰, 朱虹润, 马鸿翔. GmCHS8和GmIFS2基因共同决定大豆中异黄酮的积累. 作物学报, 2011, 37: 571-578 |
| Yi J X, Xu Z L, Wang J F, Zhang D Y, He X L, Zhu H R, Ma H X.TheGmCHS8 and GmIFS2 genes together determine the accumulation of isoflavones in soybeans. Acta Agron Sin, 2011, 37: 571-578 (in Chinese with English abstract) | |
| [11] | Butelli E, Titta L, Giorgio M, Mock H P, Matros A, Peterek S, Schijlen E G, Hall R D, Bovy A G, Luo J, Martin, C.Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat Biotechnol, 2008, 26: 1301-1308 |
| [12] | Cutanda-Perez M C, Ageorges A, Gomez C, Vialet S, Terrier N, Romieu C, Torregrosa L. Ectopic expression ofVlmybA1 in grapevine activates a narrow set of genes involved in anthocyanin synthesis and transport. Plant Mol Biol, 2009, 69: 633-648 |
| [13] | Hichri I, Barrieu F, Bogs J, Kappel C, Delrot S, Lauvergeat V.Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway.J Exp Bot, 2011, 62: 2465-2483 |
| [14] | Zhou M L, Hou H L, Zhu X M, Shao J R, Wu Y M, Tang Y X.Soybean transcription factor GmMYBZ2 represses catharanthine biosynthesis in hairy roots of Catharanthus roseus. Appl Microbiol Biotechnol, 2011, 91: 1095-1105 |
| [15] | Chen Y H, Yang X Y, He K, Liu M H, Li J G, Gao Z F, Lin Z Q, Zhang Y F, Wang X X, Qiu X M, Shen Y P, Zhang L, Deng X H, Luo J C, Deng X W, Chen Z L, Gu H Y, Qu L J.The MYB transcription factor superfamily ofArabidopsis: expression analysis and phylogenetic comparison with the rice MYB family. Plant Mol Biol, 2006, 60: 107-124 |
| [16] | Jin H, Martin C.Multifunctionality and diversity within the plant MYB-gene family.Plant Mol Biol, 1999, 41: 577-585 |
| [17] | Stracke R, Werber M, Weisshaar B.The R2R3-MYB gene family in Arabidopsis thaliana. Curr Opin Plant Biol, 2001, 4: 447-456 |
| [18] | Du H, Yang S S, Liang Z, Feng B R, Liu L, Huang Y B, Tang Y X.Genome-wide analysis of the MYB transcription factor superfamily in soybean.BMC Plant Biol, 2012, 12: 106 |
| [19] | Chu S, Wang J, Zhu Y, Liu S, Zhou X, Zhang H, Wang C E, Yang W, Tian Z, Cheng H, Yu D Y.An R2R3-type MYB transcription factor, GmMYB29, regulates isoflavone biosynthesis in soybean.PLoS Genet, 2017, 13: e1006770 |
| [20] | Liu X, Yuan L, Xu L, Xu Z, Huang Y, He X, Ma H, Yi J, Zhang D.Over-expression ofGmMYB39 leads to an inhibition of the isoflavonoid biosynthesis in soybean(Glycine max L.). Plant Biotechnol Rep, 2013, 7: 445-455 |
| [21] | Yan J, Wang B, Zhong Y, Yao L, Cheng L, Wu T.The soybean R2R3 MYB transcription factorGmMYB100 negatively regulates plant flavonoid biosynthesis. Plant Mol Biol, 2015, 89: 35-48 |
| [22] | Robbins M P, Hartnoll J, Morris P.Phenylpropanoid defence responses in transgenicLotus corniculatus 1. Glutathione elicitation of isoflavan phytoalexins in transformed root cultures. Plant Cell Rep, 1991, 10: 59-62 |
| [23] | Shelton D, Stranne M, Mikkelsen L, Pakseresht N, Welham T, Hiraka H, Tabata S, Sato S, Paquette S, Wang T L, Martin C, Bailey P.Transcription factors of Lotus: regulation of isoflavonoid biosynthesis requires coordinated changes in transcription factor activity.Plant Physiol, 2012, 159: 531-547 |
| [24] | Dubos C, Stracke R, Grotewold E, Weisshaar B, Martin C, Lepiniec L.MYB transcription factors in Arabidopsis.Trends Plant Sci, 2010, 15: 573-581 |
| [25] | Higgins D G, Thompson J D, Gibson T J.Using CLUSTAL for multiple sequence alignments.Method Enzymol, 1996, 266: 383-402 |
| [26] | Tamura K, Stecher G, Peterson D, Filipski A, Kumar S.MEGA6: Molecular Evolutionary Genetics Analysis version 6.0.Mol Biol Evol, 2013, 30: 2725-2729 |
| [27] | Livak K J, Schmittgen T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method.Methods, 2001, 25: 402-408 |
| [28] | Sun J M, Sun B L, Han F X, Yan S R, Yang H, Kikuchi A.Rapid HPLC method for determination of 12 isoflavone components in soybean seeds.Agric Sci China, 2011, 10: 70-77 |
| [29] | Chung E S, Kim K M, Lee J H.Molecular cloning and characterization of a soybeanGmMBY184 induced by abiotic stresses. J Plant Biotechnol, 2012, 39: 175-181 |
| [30] | Ishida T, Kurata T, Okada K, Wada T.A genetic regulatory network in the development of trichomes and root hairs.Annu Review Plant Biol, 2008, 59: 365-386 |
| [31] | Nadeau J A.Stomatal development: new signals and fate determinants.Curr Opin Plant Biol, 2009, 12: 29-35 |
| [32] | Wang X, Niu Q W, Teng C, Li C, Mu J, Chua N H, Zuo J.Overexpression ofPGA37/MYB118 and MYB115 promotes vegetative-to-embryonic transition in Arabidopsis. Cell Res, 2009, 19: 224-235 |
| [33] | Muller D, Schmitz G, Theres K.Blind homologous R2R3 Myb genes control the pattern of lateral meristem initiation in Arabidopsis. Plant Cell, 2006, 18: 586-597 |
| [34] | Mu R L, Cao Y R, Liu Y F, Lei G, Zou H F, Liao Y, Wang H W, Zhang W K, Ma B, Du J Z, Yuan M, Zhang J S, Chen S Y.An R2R3-type transcription factor geneAtMYB59 regulates root growth and cell cycle progression in Arabidopsis. Cell Res, 2009, 19: 1291-1304 |
| [35] | Abe H, Yamaguchi-Shinozaki K, Urao T, Iwasaki T, Hosokawa D, Shinozaki K.Role of Arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression.Plant Cell, 1997, 9: 1859-1868 |
| [36] | Mengiste T, Chen X, Salmeron J, Dietrich R.TheBOTRYTIS SUSCEPTIBLE1 gene encodes an R2R3MYB transcription factor protein that is required for biotic and abiotic stress responses in Arabidopsis. Plant Cell, 2003, 15: 2551-2565 |
| [37] | Borevitz J O, Xia Y, Blount J, Dixon R A, Lamb C.Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis.Plant Cell, 2000, 12: 2383-2394 |
| [38] | Sablowski R W, Moyano E, Culianez-Macia F A, Schuch W, Martin C, Bevan M. A flower-specific Myb protein activates transcription of phenylpropanoid biosynthetic genes.EMBO J, 1994, 13: 128-137 |
| [39] | Solano R, Nieto C, Avila J, Canas L, Diaz I, Paz-Ares J.Dual DNA binding specificity of a petal epidermis-specific MYB transcription factor (MYB.Ph3) fromPetunia hybrida. EMBO J, 1995, 14: 1773-1784 |
| [40] | Yu O, Shi J, Hession A O, Maxwell C A, McGonigle B, Odell J T. Metabolic engineering to increase isoflavone biosynthesis in soybean seed.Phytochemistry, 2003, 63: 753-763 |
| [41] | Grotewold E, Chamberlin M, Snook M, Siame B, Butler L, Swenson J, Maddock S, St Clair G, Bowen B.Engineering secondary metabolism in maize cells by ectopic expression of transcription factors.Plant Cell, 1998, 10: 721-740 |
| [42] | Holl J, Vannozzi A, Czemmel S, D'Onofrio C, Walker A R, Rausch T, Lucchin M, Boss P K, Dry I B, Bogs J. The R2R3-MYB transcription factors MYB14 and MYB15 regulate stilbene biosynthesis inVitis vinifera. Plant Cell, 2013, 25: 4135-4149 |
| [43] | Dhaubhadel S, Gijzen M, Moy P, Farhangkhoee M.Transcriptome analysis reveals a critical role ofCHS7 and CHS8 genes for isoflavonoid synthesis in soybean seeds. Plant Physiol, 2007, 143: 326-338 |
| [1] | 陈玲玲, 李战, 刘亭萱, 谷勇哲, 宋健, 王俊, 邱丽娟. 基于783份大豆种质资源的叶柄夹角全基因组关联分析[J]. 作物学报, 2022, 48(6): 1333-1345. |
| [2] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [3] | 王炫栋, 杨孙玉悦, 高润杰, 余俊杰, 郑丹沛, 倪峰, 蒋冬花. 拮抗大豆斑疹病菌放线菌菌株的筛选和促生作用及防效研究[J]. 作物学报, 2022, 48(6): 1546-1557. |
| [4] | 于春淼, 张勇, 王好让, 杨兴勇, 董全中, 薛红, 张明明, 李微微, 王磊, 胡凯凤, 谷勇哲, 邱丽娟. 栽培大豆×半野生大豆高密度遗传图谱构建及株高QTL定位[J]. 作物学报, 2022, 48(5): 1091-1102. |
| [5] | 李阿立, 冯雅楠, 李萍, 张东升, 宗毓铮, 林文, 郝兴宇. 大豆叶片响应CO2浓度升高、干旱及其交互作用的转录组分析[J]. 作物学报, 2022, 48(5): 1103-1118. |
| [6] | 朱峥, 王田幸子, 陈悦, 刘玉晴, 燕高伟, 徐珊, 马金姣, 窦世娟, 李莉云, 刘国振. 水稻转录因子WRKY68在Xa21介导的抗白叶枯病反应中发挥正调控作用[J]. 作物学报, 2022, 48(5): 1129-1140. |
| [7] | 彭西红, 陈平, 杜青, 杨雪丽, 任俊波, 郑本川, 罗凯, 谢琛, 雷鹿, 雍太文, 杨文钰. 减量施氮对带状套作大豆土壤通气环境及结瘤固氮的影响[J]. 作物学报, 2022, 48(5): 1199-1209. |
| [8] | 陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展[J]. 作物学报, 2022, 48(4): 781-790. |
| [9] | 王好让, 张勇, 于春淼, 董全中, 李微微, 胡凯凤, 张明明, 薛红, 杨梦平, 宋继玲, 王磊, 杨兴勇, 邱丽娟. 大豆突变体ygl2黄绿叶基因的精细定位[J]. 作物学报, 2022, 48(4): 791-800. |
| [10] | 李瑞东, 尹阳阳, 宋雯雯, 武婷婷, 孙石, 韩天富, 徐彩龙, 吴存祥, 胡水秀. 增密对不同分枝类型大豆品种同化物积累和产量的影响[J]. 作物学报, 2022, 48(4): 942-951. |
| [11] | 杜浩, 程玉汉, 李泰, 侯智红, 黎永力, 南海洋, 董利东, 刘宝辉, 程群. 利用Ln位点进行分子设计提高大豆单荚粒数[J]. 作物学报, 2022, 48(3): 565-571. |
| [12] | 周悦, 赵志华, 张宏宁, 孔佑宾. 大豆紫色酸性磷酸酶基因GmPAP14启动子克隆与功能分析[J]. 作物学报, 2022, 48(3): 590-596. |
| [13] | 王娟, 张彦威, 焦铸锦, 刘盼盼, 常玮. 利用PyBSASeq算法挖掘大豆百粒重相关位点与候选基因[J]. 作物学报, 2022, 48(3): 635-643. |
| [14] | 董衍坤, 黄定全, 高震, 陈栩. 大豆PIN-Like (PILS)基因家族的鉴定、表达分析及在根瘤共生固氮过程中的功能[J]. 作物学报, 2022, 48(2): 353-366. |
| [15] | 张国伟, 李凯, 李思嘉, 王晓婧, 杨长琴, 刘瑞显. 减库对大豆叶片碳代谢的影响[J]. 作物学报, 2022, 48(2): 529-537. |
|
||