欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (2): 285-293.doi: 10.3724/SP.J.1006.2021.03015
蒋成功1,2, 石慧敏2, 王红武2, 李坤2, 黄长玲2, 刘志芳2, 吴宇锦2, 李树强2, 胡小娇2,*, 马庆1,*
JIANG Cheng-Gong1,2, SHI Hui-Min2, WANG Hong-Wu2, LI Kun2, HUANG Chang-Ling2, LIU Zhi-Fang2, WU Yu-Jin2, LI Shu-Qiang2, HU Xiao-Jiao2,*, MA Qing1,*
摘要:
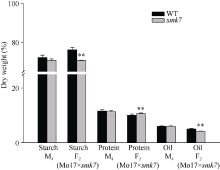
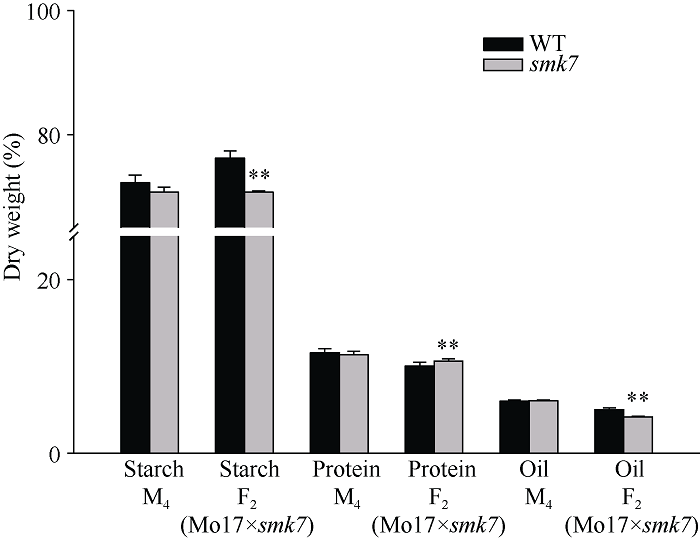
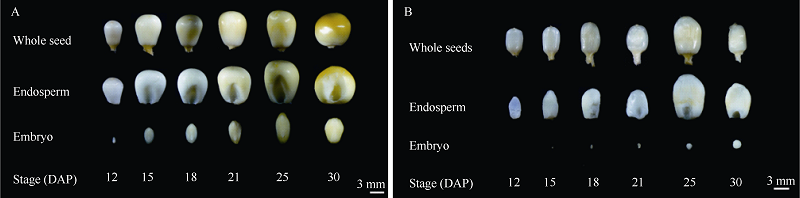
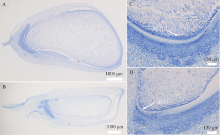
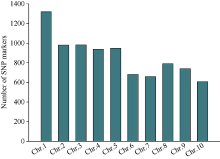
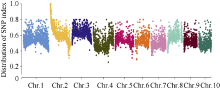
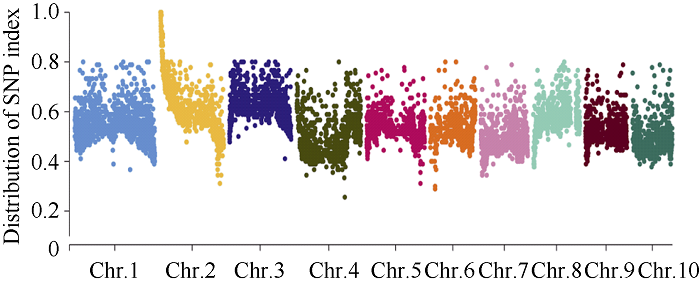
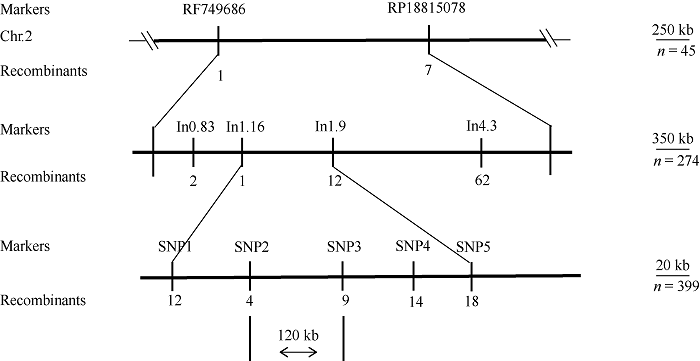
利用甲基磺酸乙酯(EMS)对玉米自交系B73进行诱变, 获得一个可以稳定遗传的小籽粒突变体smk7 (small kernel 7)。smk7成熟籽粒表现为体积变小, 胚和胚乳发育缺陷, 百粒重显著降低。突变籽粒发芽率仅为10%, 且幼苗黄化不能生长成正常植株。成熟smk7胚乳中淀粉、蛋白、油分含量与野生型籽粒相比无显著差异, 但突变体胚乳淀粉粒体积明显变小且形状不规则。smk7突变籽粒在授粉后12 d即可观察到明显的小籽粒和空瘪表型, 石蜡切片显微观察显示突变籽粒的胚和胚乳发育迟缓, 胚乳基部转移层细胞(BETL)相对于野生型细胞壁向内生长减少, 发育受阻。用杂合植株(+/smk7)与多个自交系分别杂交, 构建不同背景的F2分离群体, 遗传分析结果表明该性状受单隐性核基因控制。利用靶向测序基因型分型(genotyping by target sequencing, GBTS)技术将基因初定位于2号染色体短臂, 进一步精细定位发现该基因位于RM1433917和RM1535316两个标记之间约120 kb的物理范围内, 共有8个蛋白编码基因。本研究为进一步克隆和解析SMK7基因调控玉米籽粒发育的分子机制奠定了基础。
| [1] |
Nuss E T, Tanumihardjo S A. Quality protein maize for Africa: closing the protein inadequacy gap in vulnerable populations. Adv Nutr, 2011,2:217-224.
doi: 10.3945/an.110.000182 pmid: 22332054 |
| [2] |
Berger F. Endosperm development. Curr Opin Plant Biol, 1999,2:28-32.
doi: 10.1016/s1369-5266(99)80006-5 pmid: 10047564 |
| [3] | Guo Z F, Wang , Tao J J, Ren Y H, Xu C, Wu K S, Zou C, Zhang J A, Xu Y B. Development of multiple SNP marker panels affordable to breeders through genotyping by target sequencing (GBTS) in maize. Mol Breed, 2019,39:37-49. |
| [4] |
Sun X, Shantharaj D, Kang X, Ni M. Transcriptional and hormonal signaling control of Arabidopsis seed development. Curr Opin Plant Biol, 2010,13:611-620.
doi: 10.1016/j.pbi.2010.08.009 pmid: 20875768 |
| [5] | 任雪梅. 玉米籽粒发育基因Emp11及ZmSmk3的克隆及功能分析. 华中农业大学博士学位论文, 湖北武汉, 2018. |
| Ren X M. Cloning and Functional Analysis of Emp11 and ZmSmk3 Controlling Kernel Development in Maize PhD Dissertation of Huazhong Agricultural University, Wuhan, Hubei, China, 2018 (in Chinese with English abstract). | |
| [6] | 李见坤. 玉米籽粒发育基因UBL1的克隆与功能分析. 中国农业大学博士学位论文, 北京, 2016. |
| Li J K. Cloning and Characterization of UBL1 Controlling Kernel Development in Maize PhD Dissertation of China Agricultural University, Beijing, China, 2016 (in Chinese with English abstract). | |
| [7] |
Neuffer M G, Sheridan W F. Defective kernel mutants of maize: I. Genetic and lethality studies. Genetics, 1980,95:929-944.
pmid: 17249053 |
| [8] |
Wang G, Sun X, Wang G, Wang F, Gao Q, Sun X, Tang Y, Chang C, Lai J, Zhu L, Xu Z, Song R. Opaque7 encodes an acyl-activating enzyme-like protein that affects storage protein synthesis in maize endosperm. Genetics, 2011,189:1281-1295.
doi: 10.1534/genetics.111.133967 |
| [9] | 姚东升. 玉米opaque10突变体基因的图位克隆和功能分析. 上海大学博士学位论文, 上海, 2016. |
| Yao D S. The Map-based Cloning and Functional Analysis of Maize opaque10 Gene PhD Dissertation of Shanghai University, Shanghai, China, 2016 (in Chinese with English abstract). | |
| [10] |
Yao D, Qi W, Li X, Yang Q, Yan S, Ling H, Wang G, Wang G, Song R. Maize opaque10 encodes a cereal-specific protein that is essential for the proper distribution of zeins in endosperm protein bodies. PLoS Genet, 2016,12:e1006270.
doi: 10.1371/journal.pgen.1006270 pmid: 27541862 |
| [11] |
Holding D R, Otegui M S, Li B, Meeley R B, Dam T, Hunter B G, Jung R, Larkins B A, Larkins B A. The maize floury1 gene encodes a novel endoplasmic reticulum protein involved in zein protein body formation. Plant Cell, 2007,19:2569-2582.
doi: 10.1105/tpc.107.053538 pmid: 17693529 |
| [12] |
Mertz E T, Bates L S, Nelson O E. Mutant gene that changes protein composition and increases lysine content of maize endosperm. Science, 1964,145:279-280.
pmid: 14171571 |
| [13] |
Schmidt R J, Burr F A, Aukerman M J, Burr B. Maize regulatory gene opaque-2 encodes a protein with a“leucine-zipper”motif that binds to zein DNA. Proc Natl Acad Sci USA, 1990,87:46-50.
doi: 10.1073/pnas.87.1.46 pmid: 2296602 |
| [14] |
Schmidt R J, Ketudat M, Aukerman M J, Hoschek G. Opaque-2 is a transcriptional activator that recognizes a specific target site in 22-kD zein genes. Plant Cell, 1992,4:689-700.
doi: 10.1105/tpc.4.6.689 pmid: 1392590 |
| [15] |
Cord-Neto G, Yunes J A, da Silva M J, Vettore A L, Arruda P, Leite A. The involvement of opaque 2 on beta-prolamin gene regulation in maize and coix suggests a more general role for this transcriptional activator. Plant Mol Biol, 1995,27:1015-1029.
doi: 10.1007/BF00037028 pmid: 7766871 |
| [16] |
Li Q, Wang J, Ye J, Zheng X, Xiang X, Li C, Fu M, Wang Q, Zhang Z, Wu Y. The maize imprinted gene FLOURY3 encodes a PLATZ protein required for tRNA and 5S rRNA transcription through interaction with RNA polymerase III. Plant Cell, 2017,29:2661-2675.
doi: 10.1105/tpc.17.00576 pmid: 28874509 |
| [17] |
Silva-Sanchez C, Chen S, Li J, Chourey P S. A comparative glycoproteome study of developing endosperm in the hexose-deficient miniature1 (mn1) seed mutant and its wild type Mn1 in maize. Front Plant Sci, 2014,5:63-77.
doi: 10.3389/fpls.2014.00063 pmid: 24616729 |
| [18] |
Zhu C, Jin G, Fang P, Zhang Y, Feng X, Tang Y, Qi W, Song R. Maize pentatricopeptide repeat protein DEK41 affects cis-splicing of mitochondrial nad4 intron 3 and is required for normal seed development. J Exp Bot, 2019,70:3795-3808.
doi: 10.1093/jxb/erz193 pmid: 31020318 |
| [19] |
Tian Q, Olsen L, Sun B, Lid S E, Brown R C, Lemmon B E, Fosnes K, Gruis D F, Opsahl-Sorteberg H G, Otegui M S, Olsen O A. Subcellular localization and functional domain studies of DEFECTIVE KERNEL1 in maize and Arabidopsis suggest a model for aleurone cell fate specification involving CRINKLY4 and SUPERNUMERARY ALEURONE LAYER1. Plant Cell, 2007,19:3127-3145.
pmid: 17933905 |
| [20] |
Yuan N, Wang J, Zhou Y, An D, Xiao Q, Wang W, Wu Y. EMB-7L is required for embryogenesis and plant development in maize involved in RNA splicing of multiple chloroplast genes. Plant Sci, 2019,287:110203.
doi: 10.1016/j.plantsci.2019.110203 pmid: 31481208 |
| [21] |
Shen Y, Li C, McCarty D R, Meeley R, Tan B C. Embryo defective12 encodes the plastid initiation factor 3 and is essential for embryogenesis in maize. Plant J, 2013,74:792-804.
doi: 10.1111/tpj.12161 pmid: 23451851 |
| [22] |
Ding S, Liu X Y, Wang H C, Wang Y, Tang J J, Yang Y Z, Tan B C. SMK6 mediates the C-to-U editing at multiple sites in maize mitochondria. J Plant Physiol, 2019,240:152992.
pmid: 31234031 |
| [23] |
Wang H C, Sayyed A, Liu X Y, Yang Y Z, Sun F, Wang Y, Wang M, Tan B C. SMALL KERNEL4 is required for mitochondrial cox1 transcript editing and seed development in maize. J Integr Plant Biol, 2019.
pmid: 33215867 |
| [24] |
Lurin C, Andres C, Aubourg S, Bellaoui M, Bitton F, Bruyere C, Caboche M, Debast C, Gualberto J, Hoffmann B, Lecharny A, Le Ret M, Martin-Magniette M L, Mireau H, Peeters N, Renou J P, Szurek B, Taconnat L, Small I. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell, 2004,16:2089-2103.
doi: 10.1105/tpc.104.022236 pmid: 15269332 |
| [25] |
Wang Y, Liu W, Wang H, Du Q, Fu Z, Li W X, Tang J. ZmEHD1 is required for kernel development and vegetative growth through regulating auxin homeostasis. Plant Physiol, 2019,182:1467-1480.
doi: 10.1104/pp.19.01336 pmid: 31857426 |
| [26] |
Li X J, Zhang Y F, Hou M, Sun F, Shen Y, Xiu Z H, Wang X, Chen Z L, Sun S S, Small I, Tan B C. Small kernel 1 encodes a pentatricopeptide repeat protein required for mitochondrial nad7 transcript editing and seed development in maize (Zea mays) and rice (Oryza sativa). Plant J, 2014,79:797-809.
doi: 10.1111/tpj.12584 pmid: 24923534 |
| [27] |
Yang Y Z, Ding S, Wang Y, Li C L, Shen Y, Meeley R, McCarty D R, Tan B C. Small kernel2 encodes a glutaminase in Vitamin B6 biosynthesis essential for maize seed development. Plant Physiol, 2017,174:1127-1138.
pmid: 28408540 |
| [28] | Pan Z, Ren X, Zhao H, Liu L, Tan Z, Qiu F. A mitochondrial transcription termination factor, ZmSmk3, is required for nad1 intron4 and nad4 intron1 splicing and kernel development in maize. G3: Genes Genom Genet, 2019,9:2677-2686. |
| [29] |
Pan Z, Liu M, Xiao Z, Ren X, Zhao H, Gong D, Liang K, Tan Z, Shao Y, Qiu F. ZmSMK9, a pentatricopeptide repeat protein, is involved in the cis-splicing of nad5, kernel development and plant architecture in maize. Plant Sci, 2019,288:110205.
doi: 10.1016/j.plantsci.2019.110205 pmid: 31521217 |
| [30] |
Handa N, Terada T, Kamewari Y, Hamana H, Tame J R, Park S Y, Kinoshita K, Ota M, Nakamura H, Kuramitsu S, Shirouzu M, Yokoyama S. Crystal structure of the conserved protein TT1542 from thermus thermophilus HB8. Protein Sci, 2003,12:1621-1632.
doi: 10.1110/gad.03104003 pmid: 12876312 |
| [31] | 王家利, 刘冬成, 郭小丽, 张爱民. 生长素合成途径的研究进展. 植物学报, 2012,47:292-301. |
| Wang J L, Liu D C, Guo X L, Zhang A M. Research progress of auxin synthesis pathway. Chin Bull Bot, 2012,47:292-301 (in Chinese with English abstract). | |
| [32] |
Bashline L, Li S, Zhu X, Gu Y. The TWD40-2 protein and the AP2 complex cooperate in the clathrin-mediated endocytosis of cellulose synthase to regulate cellulose biosynthesis. Proc Natl Acad Sci USA, 2015,112:12870-12875.
doi: 10.1073/pnas.1509292112 pmid: 26417106 |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 郑崇珂, 周冠华, 牛淑琳, 和亚男, 孙伟, 谢先芝. 水稻早衰突变体esl-H5的表型鉴定与基因定位[J]. 作物学报, 2022, 48(6): 1389-1400. |
| [4] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [5] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [6] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [7] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [8] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [9] | 王好让, 张勇, 于春淼, 董全中, 李微微, 胡凯凤, 张明明, 薛红, 杨梦平, 宋继玲, 王磊, 杨兴勇, 邱丽娟. 大豆突变体ygl2黄绿叶基因的精细定位[J]. 作物学报, 2022, 48(4): 791-800. |
| [10] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [11] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [12] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [13] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [14] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [15] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
|
||