欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (4): 812-824.doi: 10.3724/SP.J.1006.2022.14076
徐昕( ), 秦超(), 赵涛, 刘斌, 李宏宇*(), 刘军*()
), 秦超(), 赵涛, 刘斌, 李宏宇*(), 刘军*()
XU Xin(), QIN Chao(), ZHAO Tao, LIU Bin, LI Hong-Yu*(), LIU Jun*()
摘要:
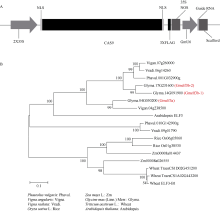
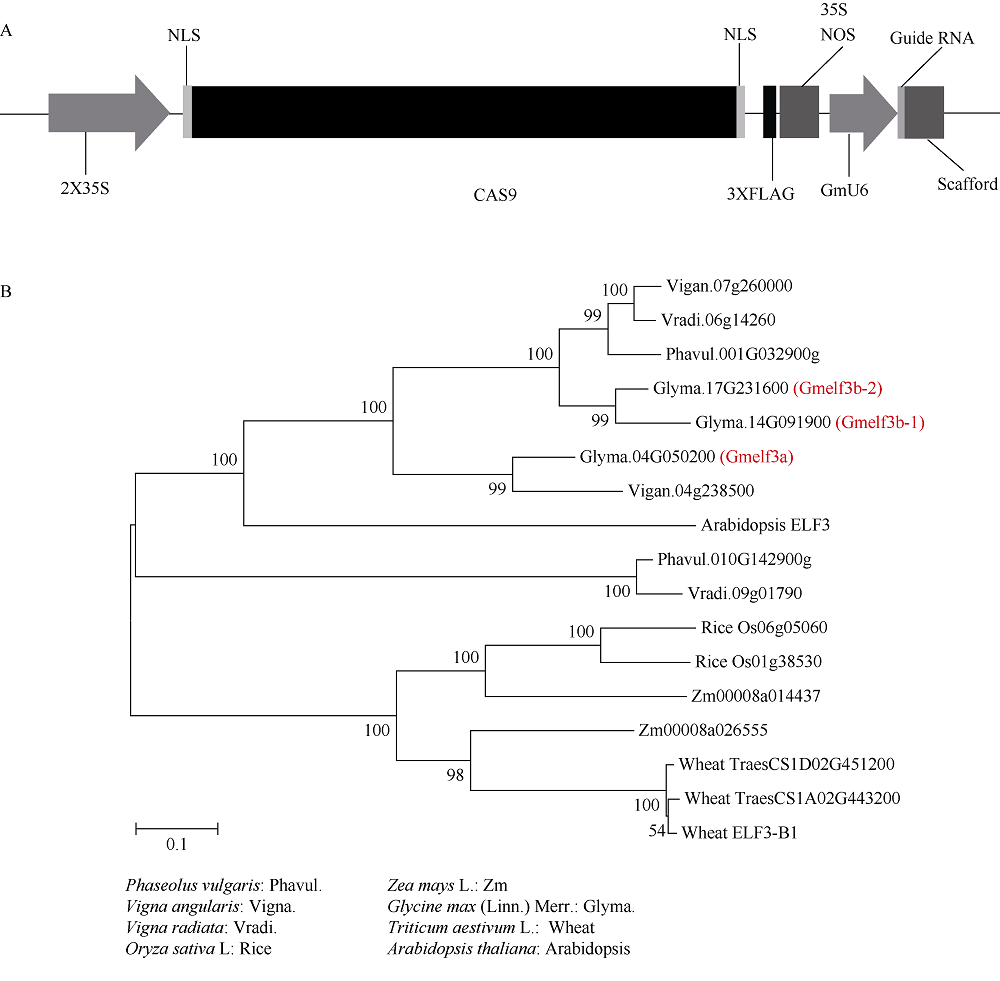
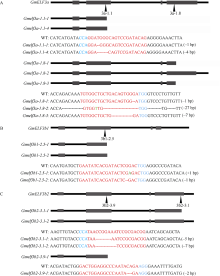
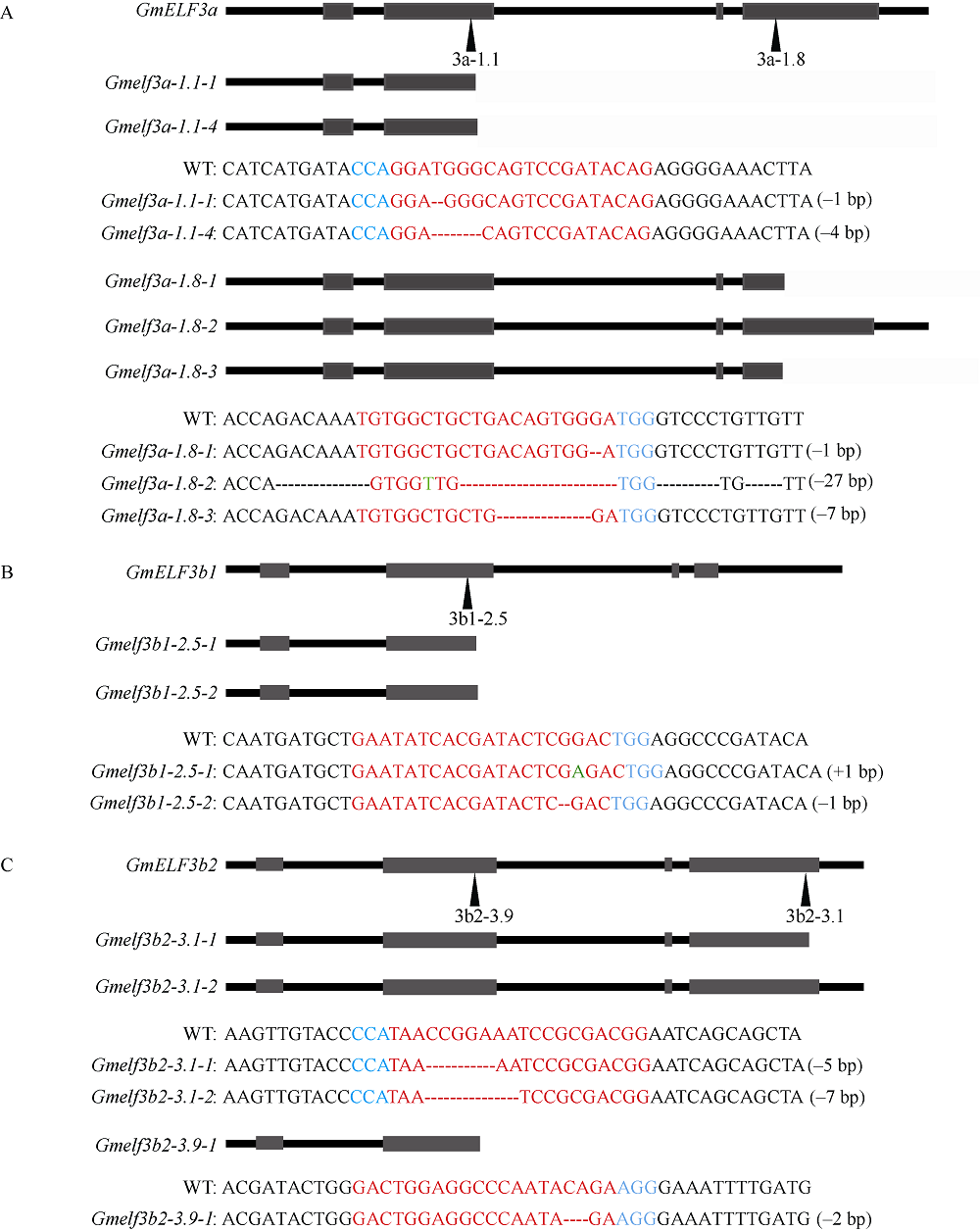
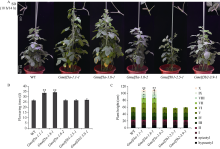
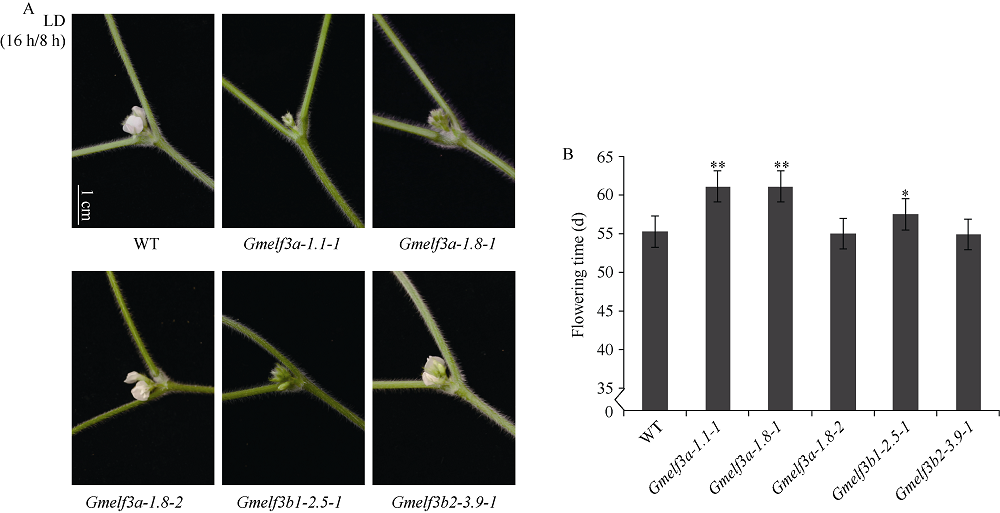
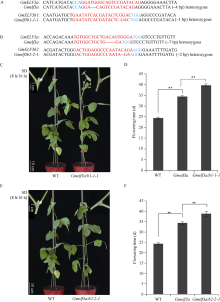
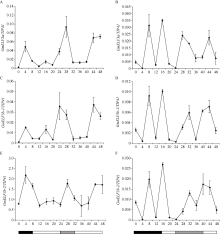
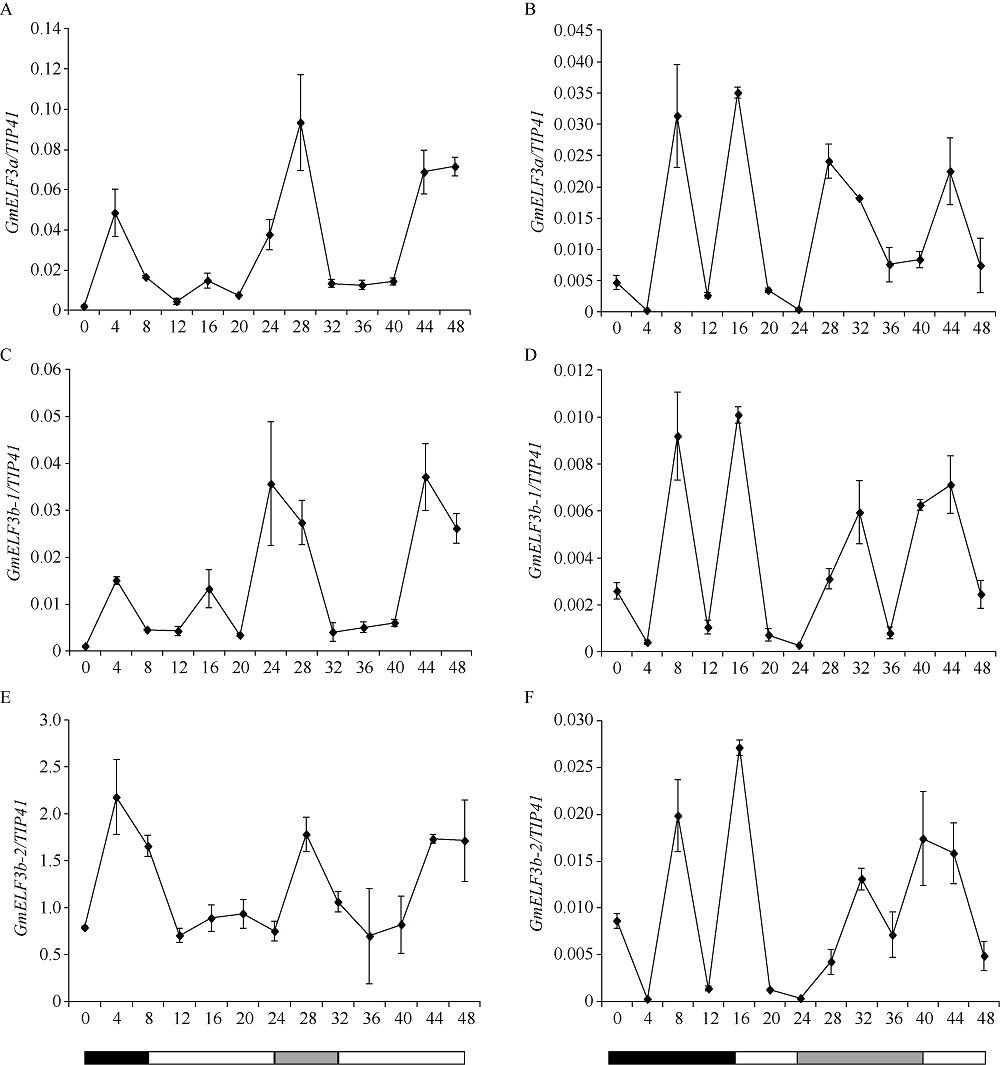
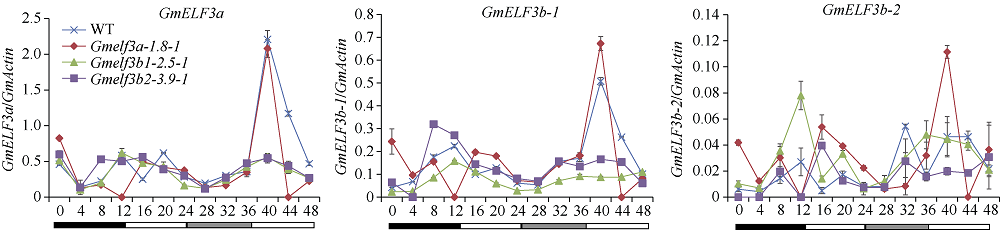
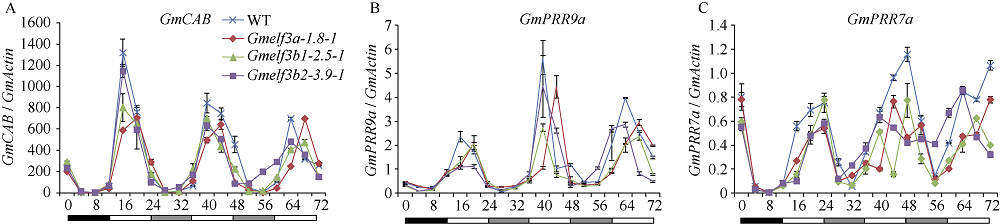
大豆是典型的短日照作物, 光周期的敏感性严重影响大豆的开花时间和产量, 制约大豆的种植范围, 但调控大豆光周期和生物钟节律的机制尚不十分清楚。在模式植物拟南芥中, ELF3与ELF4、LUX一起, 形成ELF4-ELF3-LUX (Evening Complex, EC)生物钟晚间复合物, 在生物钟节律和开花时间调控等方面发挥重要作用。本研究通过CRISPR/Cas9基因编辑系统获得大豆Gmelf3a/j、Gmelf3b-1和Gmelf3b-2的突变体材料。通过观察Gmelf3a/j、Gmelf3b-1和Gmelf3b-2各突变体材料在短日照和长日照下的开花时间发现, GmELF3b-1在长日照下对大豆开花时间起调控作用; 通过观察非纯合双突变体的表型发现, GmELF3a/J与GmELF3b-1和GmELF3b-2之间在调控大豆开花时间方面存在功能冗余。通过qRT-PCR对大豆生物钟节律相关基因的表达进行检测发现, GmCAB、GmPRR9a和GmPRR7a的表达模式发生改变, 这表明GmELF3a/J、GmELF3b-1和GmELF3b-2可能是通过GmPRR9a和GmPRR7a对大豆生物钟节律和开花时间进行调控。
| [1] |
Bernard R L. Two major genes for time of flowering and maturity in soybeans. Crop Sci, 1971, 11:242-244.
doi: 10.2135/cropsci1971.0011183X001100020022x |
| [2] |
Buzzell R I. Inheritance of a soybean flowering response to fluorescent-daylength conditions. Can J Genet Cytol, 1971, 13:703-707.
doi: 10.1139/g71-100 |
| [3] |
Watanabe S, Harada K, Abe J. Genetic and molecular bases of photoperiod responses of flowering in soybean. Breed Sci, 2012, 61:531-543.
doi: 10.1270/jsbbs.61.531 |
| [4] |
Hartwig E E, Kiihl R A S. Identification and utilization of a delayed flowering character in soybeans for short-day conditions. Field Crops Res, 1979, 2:145-151.
doi: 10.1016/0378-4290(79)90017-0 |
| [5] |
Sinclair T R, Hinson K. Soybean flowering in response to the long-juvenile trait. Crop Sci, 1992, 32:1242-1248.
doi: 10.2135/cropsci1992.0011183X003200050036x |
| [6] |
Carpentieri-Pípolo V, Almeida L A, Kiihl R A S. Inheritance of a long juvenile period under short-day conditions in soybean. Genet Mol Biol, 2002, 25:463-469.
doi: 10.1590/S1415-47572002000400016 |
| [7] |
Lu S J, Zhao X H, Hu Y L, Liu S L, Nan H Y, Li X M, Fang C, Cao D, Shi X Y, Kong L P, Su T, Zhang F G, Li S C, Wang Z, Yuan X H, Cober E R, Weller J L, Liu B H, Hou X L, Tian Z X, Kong F J. Natural variation at the soybean J locus improves adaptation to the tropics and enhances yield. Nat Genet, 2017, 49:773-779.
doi: 10.1038/ng.3819 |
| [8] |
Yue Y L, Liu N X, Jiang B J, Li M, Wang H J, Jiang Z, Pan H T, Xia Q J, Ma Q B, Han T F, Nian H. A single nucleotide deletion in J encoding GmELF3 confers long juvenility and is associated with adaption of tropic soybean. Mol Plant, 2017, 10:656-658.
doi: 10.1016/j.molp.2016.12.004 |
| [9] |
Cheng Q, Gan Z R, Wang Y P, Lu S J, Hou Z H, Li H Y, Xiang H T, Liu B H, Kong F J, Dong L D. The soybean gene J contributes to salt stress tolerance by up-regulating salt-responsive genes. Front Plant Sci, 2020, 11:272.
doi: 10.3389/fpls.2020.00272 pmid: 32256507 |
| [10] |
Fang C, Liu J, Zhang T, Su T, Li S C, Cheng Q, Kong L P, Li X M, Bu T T, Li H Y, Dong L D, Lu S J, Kong F J, Liu B H. A recent retrotransposon insertion of J caused E6 locus facilitating soybean adaptation into low latitude. J Integr Plant Biol, 2021, 63:995-1003.
doi: 10.1111/jipb.v63.6 |
| [11] |
Inoue K, Araki T, Endo M. Circadian clock during plant development. J Plant Res, 2018, 131:59-66.
doi: 10.1007/s10265-017-0991-8 |
| [12] |
Johansson M, Staiger D. Time to flower: interplay between photoperiod and the circadian clock. J Exp Bot, 2015, 66:719-730.
doi: 10.1093/jxb/eru441 pmid: 25371508 |
| [13] |
Hicks K A, Albertson T M, Wagner D R. EARLY FLOWERING3 encodes a novel protein that regulates circadian clock function and flowering in Arabidopsis. Plant Cell, 2001, 13:1281-1292.
pmid: 11402160 |
| [14] |
Liu X L, Covington M F, Fankhauser C, Chory J, Wagner D R. ELF3 encodes a circadian clock-regulated nuclear protein that functions in an Arabidopsis PHYB signal transduction pathway. Plant Cell, 2001, 13:1293-1304.
pmid: 11402161 |
| [15] |
Doyle M R, Davi S J, Bastow R M, McWatters H G, Kozma- Bognar L, Nagy F, Milla A J, Amasin R M. The ELF4 gene controls circadian rhythms and flowering time in Arabidopsis thaliana. Nature, 2002, 419:74-77.
doi: 10.1038/nature00954 |
| [16] |
Nusinow D A, Helfer A, Hamilton E E, King J J, Imaizumi T, Schultz T F, Farre E M, Kay S A. The ELF4-ELF3-LUX complex links the circadian clock to diurnal control of hypocotyl growth. Nature, 2011, 475:398-402.
doi: 10.1038/nature10182 |
| [17] |
Liew L C, Singh M B, Bhalla P L. A novel role of the soybean clock gene LUX ARRHYTHMO in male reproductive development. Sci Rep, 2017, 7:10605.
doi: 10.1038/s41598-017-00205-9 |
| [18] |
Fang X L, Han Y P, Liu M S, Jiang J C, Li X, Lian Q C, Xie X R, Huang Y, Ma Q B, Nian H, Qi J, Yan C Y, Wang Y X. Modulation of evening complex activity enables north-to-south adaptation of soybean. Sci China Life Sci, 2021, 64:179-195.
doi: 10.1007/s11427-020-1832-2 |
| [19] |
Brambilla V, Fornara F. Molecular control of flowering in response to day length in rice. J Integr Plant Biol, 2013, 55:410-418.
doi: 10.1111/jipb.12033 |
| [20] | Hazen S P, Schultz T F, Pruneda-Paz J L, Borevitz J O, Ecker J R, Kay S A. LUX ARRHYTHMO encodes a Myb domain protein essential for circadian rhythms. Proc Natl Acad Sci USA, 2005, 102:10387-10392. |
| [21] |
Saito H, Ogiso-Tanaka E, Okumoto Y, Yoshitake Y, Izumi H, Yokoo T, Matsubara K, Hori K, Yano M, Inoue H, Tanisaka T. Ef7 encodes an ELF3-like protein and promotes rice flowering by negatively regulating the floral repressor gene Ghd7 under both short- and long-day conditions. Plant Cell Physiol, 2012, 53:717-728.
doi: 10.1093/pcp/pcs029 |
| [22] |
Alvarez M A, Tranquilli G, Lewis S, Kippes N, Dubcovsky J. Genetic and physical mapping of the earliness per se locus Eps-A (m) 1 in Triticum monococcum identifies EARLY FLOWERING 3 (ELF3) as a candidate gene. Funct Integr Genomics, 2016, 16:365-382.
doi: 10.1007/s10142-016-0490-3 |
| [23] |
Faure S, Turner A S, Gruszka D, Christodoulou V, Davis S J, Korff M V, Laurie D A. Mutation at the circadian clock gene EARLY MATURITY 8 adapts domesticated barley (Hordeum vulgare) to short growing seasons. Proc Natl Acad Sci USA, 2012, 109:8328-8333.
doi: 10.1073/pnas.1120496109 |
| [24] | Weller J L, Liew L C, Hecht V F G, Rajandran V, Laurie R E, Ridge S, Wenden B, Vander Schoor J K, Jaminon O, Blassiau C, Dalmais M, Rameau C, Bendahmane A, Macknight R C, Lejeune-Henaut I. A conserved molecular basis for photoperiod adaptation in two temperate legumes. Proc Natl Acad Sci USA, 2012, 109:21158-21163. |
| [25] |
Zakhrabekova S, Gough S P, Braumann I, Muller A H, Lundqvist J, Ahmann K, Dockter C, Matyszczak I, Kurowska M, Druka A, Waugh R, Graner A, Stein N, Steuernagel B, Lundqvist U, Hansson M. Induced mutations in circadian clock regulator Mat-a facilitated short-season adaptation and range extension in cultivated barley. Proc Natl Acad Sci USA, 2012, 109:4326-4331.
doi: 10.1073/pnas.1113009109 |
| [26] | Bu T T, Lu S J, Wang K, Dong L D, Li S L, Xie Q G, Xu X D, Cheng Q, Chen L Y, Fang C, Li H Y, Liu B H, Weller J L, Kong F J. A critical role of the soybean evening complex in the control of photoperiod sensitivity and adaptation. Proc Natl Acad Sci USA, 2021, 118:e2010241118. |
| [27] |
Pokhilko A, Fernandez A P, Edwards K D, Southern M M, Halliday K J, Millar A J. The clock gene circuit in Arabidopsis includes a repressilator with additional feedback loops. Mol Syst Biol, 2012, 8:574.
doi: 10.1038/msb.2012.6 |
| [28] |
Hsu P Y, Harmer S L. Wheels within wheels: the plant circadian system. Trends Plant Sci, 2014, 19:240-249.
doi: 10.1016/j.tplants.2013.11.007 |
| [29] |
Dixon L E, Knox K, Kozma-Bognar L, Southern M M, Pokhilko A, Millar A J. Temporal repression of core circadian genes is mediated through EARLY FLOWERING 3 in Arabidopsis. Curr Biol, 2011, 21:120-125.
doi: 10.1016/j.cub.2010.12.013 |
| [30] |
Helfer A, Nusinow D A, Chow B Y, Gehrke A R, Bulyk M L, Kay S A. LUX ARRHYTHMO encodes a nighttime repressor of circadian gene expression in the Arabidopsis core clock. Curr Biol, 2011, 21:126-133.
doi: 10.1016/j.cub.2010.12.021 pmid: 21236673 |
| [31] |
Chow B Y, Helfer A, Nusinow D A, Kay S A. ELF3 recruitment to the PRR9 promoter requires other evening complex members in the Arabidopsis circadian clock. Plant Signal Behav, 2012, 7:170-173.
doi: 10.4161/psb.18766 |
| [32] |
Herrero E, Kolmos E, Bujdoso N, Yuan Y, Wang M, Berns M C, Uhlworm H, Coupland G, Saini R, Jaskolski M, Webb A, Goncalves J, Davis S J. EARLY FLOWERING4 recruitment of EARLY FLOWERING3 in the nucleus sustains the Arabidopsis circadian clock. Plant Cell, 2012, 24:428-443.
doi: 10.1105/tpc.111.093807 |
| [33] |
Kamioka M, Takao S, Suzuki T, Taki K, Higashiyama T, Kinoshita T, Nakamichi N. Direct repression of evening genes by CIRCADIAN CLOCK-ASSOCIATED1 in the Arabidopsis circadian clock. Plant Cell, 2016, 28:696-711.
doi: 10.1105/tpc.15.00737 |
| [1] | 贾小平,袁玺垒,李剑峰,王永芳,张小梅,张博,全建章,董志平. 不同光温条件谷子光温互作模式研究及SiCCT基因表达分析[J]. 作物学报, 2020, 46(7): 1052-1062. |
| [2] | 张双双,王立伟,姚楠,郭光艳,夏玉凤,秘彩莉. 水稻OsUBA基因的表达及其在促进种子萌发和开花中的功能[J]. 作物学报, 2019, 45(9): 1327-1337. |
| [3] | 贾小平,全建章,王永芳,董志平,袁玺垒,张博,李剑峰. 不同光周期环境对谷子农艺性状的影响[J]. 作物学报, 2019, 45(7): 1119-1127. |
| [4] | 马晨雨,詹为民,李文亮,张梦迪,席章营. 玉米ZmNAOD基因的克隆与功能分析[J]. 作物学报, 2018, 44(10): 1433-1441. |
| [5] | 张立武,黄枝秒,万雪贝,林荔辉,徐建堂,陶爱芬,方平平,祁建民. 红麻光周期钝感材料的鉴定与遗传分析[J]. 作物学报, 2014, 40(12): 2098-2103. |
| [6] | 夏正俊. 大豆光周期反应与生育期基因研究进展[J]. 作物学报, 2013, 39(04): 571-579. |
| [7] | 曹岩,张晓玫,陈新建,傅永福. 大豆GmNF-YC2基因的克隆与功能分析[J]. 作物学报, 2012, 38(09): 1607-1616. |
| [8] | 杨芳萍, 韩利明, 阎俊, 夏先春, 张勇, 曲延英, 王忠伟, 何中虎. 春化和光周期基因等位变异在23个国家小麦品种中的分布[J]. 作物学报, 2011, 37(11): 1917-1925. |
| [9] | 王翠玲, 孙朝辉, 库丽霞, 王铁固, 陈彦惠. 利用永久F2群体在不同光周期环境下定位玉米株高QTL[J]. 作物学报, 2011, 37(02): 271-279. |
| [10] | 库丽霞,孙朝辉,王翠玲,张君,张伟强,陈彦惠. 玉米光周期敏感相关性状发育动态QTL定位[J]. 作物学报, 2010, 36(4): 602-611. |
| [11] | 张清哲,马锦花,陈新建,傅永福. 大豆GmCOL4基因的克隆与分析[J]. 作物学报, 2010, 36(4): 539-548. |
| [12] | 曲明南, 孙石, 吴存祥, 费志宏, 郭玉华, 侯文胜, 韩天富. 对早熟、矮杆、小粒大豆基因型MiniMax作为大豆研究模式材料的探讨[J]. 作物学报, 2010, 36(11): 1990-1997. |
| [13] | 张德贵,李新海,李明顺,谢传晓,郝转芳,白丽,张世煌. 四个热带亚热带玉米群体开花期性状对混合选择的直接选择响应[J]. 作物学报, 2010, 36(1): 28-35. |
| [14] | 王英,程立锐,冷建田,吴存祥,侯文胜,韩天富. 开花后不同光周期条件下大豆农艺性状和品质性状的QTL分析[J]. 作物学报, 2010, 36(07): 1092-1099. |
| [15] | 费志宏,吴存祥,孙洪波,侯文胜,张宝石,韩天富. 以光周期处理与分期播种试验综合鉴定大豆品种的光温反应[J]. 作物学报, 2009, 35(8): 1525-1531. |
|
||