欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (3): 576-589.doi: 10.3724/SP.J.1006.2024.31025
张宝华1,2( ), 刘佳静1,2, 田晓1,2, 田旭钊1,2, 董阔1, 武郁洁1, 肖凯3,*(), 李小娟1,2,*()
), 刘佳静1,2, 田晓1,2, 田旭钊1,2, 董阔1, 武郁洁1, 肖凯3,*(), 李小娟1,2,*()
ZHANG Bao-Hua1,2(), LIU Jia-Jing1,2, TIAN Xiao1,2, TIAN Xu-Zhao1,2, DONG Kuo1, WU Yu-Jie1, XIAO Kai3,*(), LI Xiao-Juan1,2,*()
摘要:
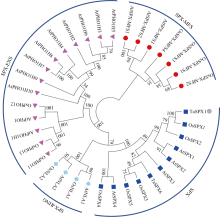
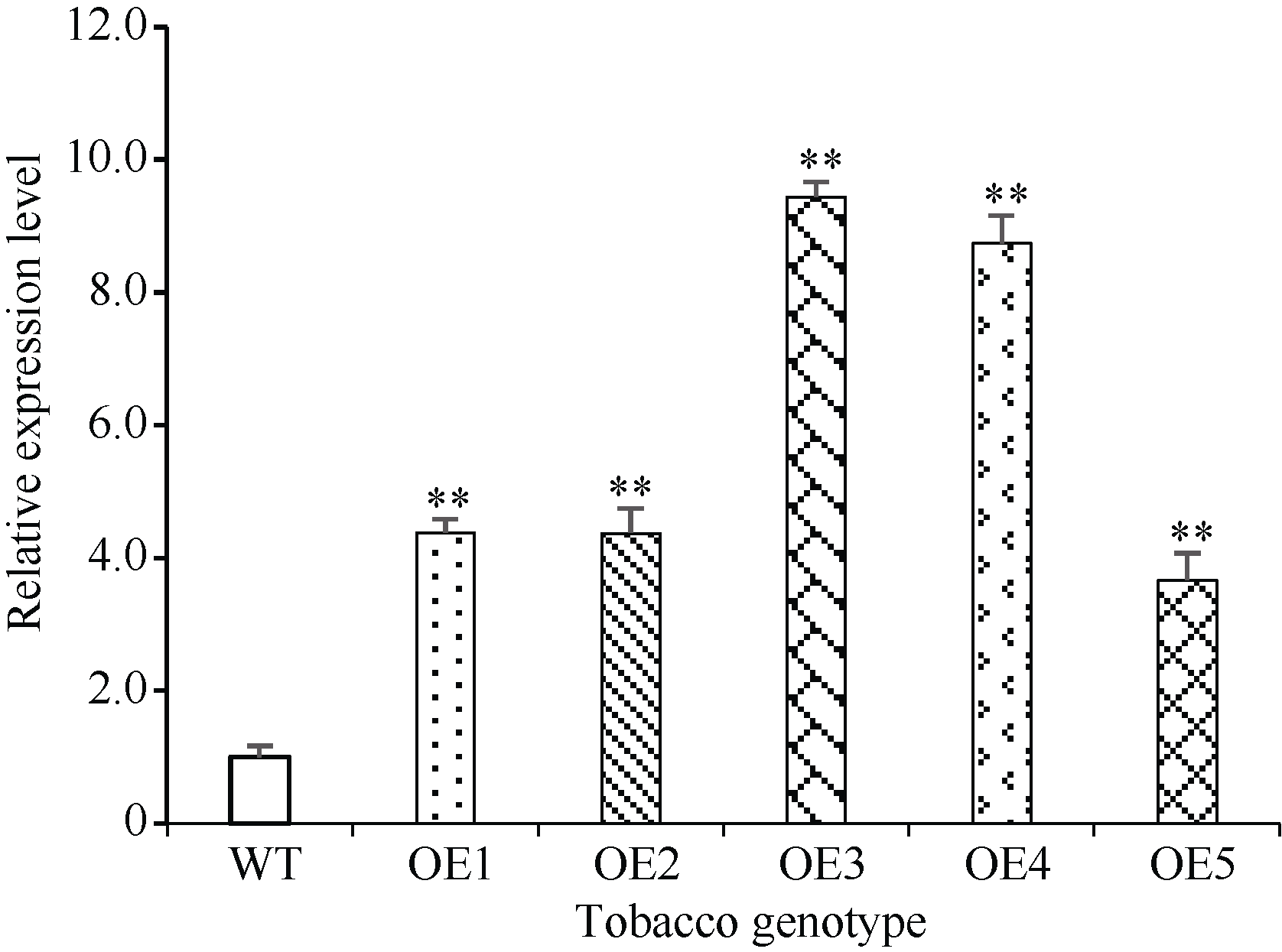
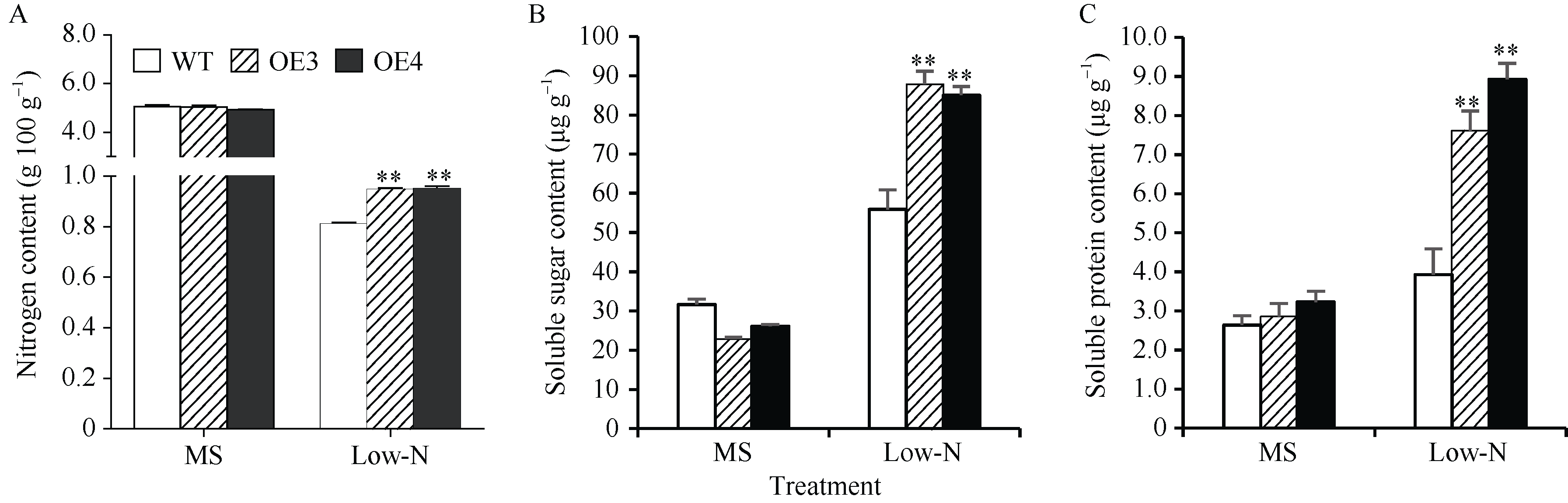
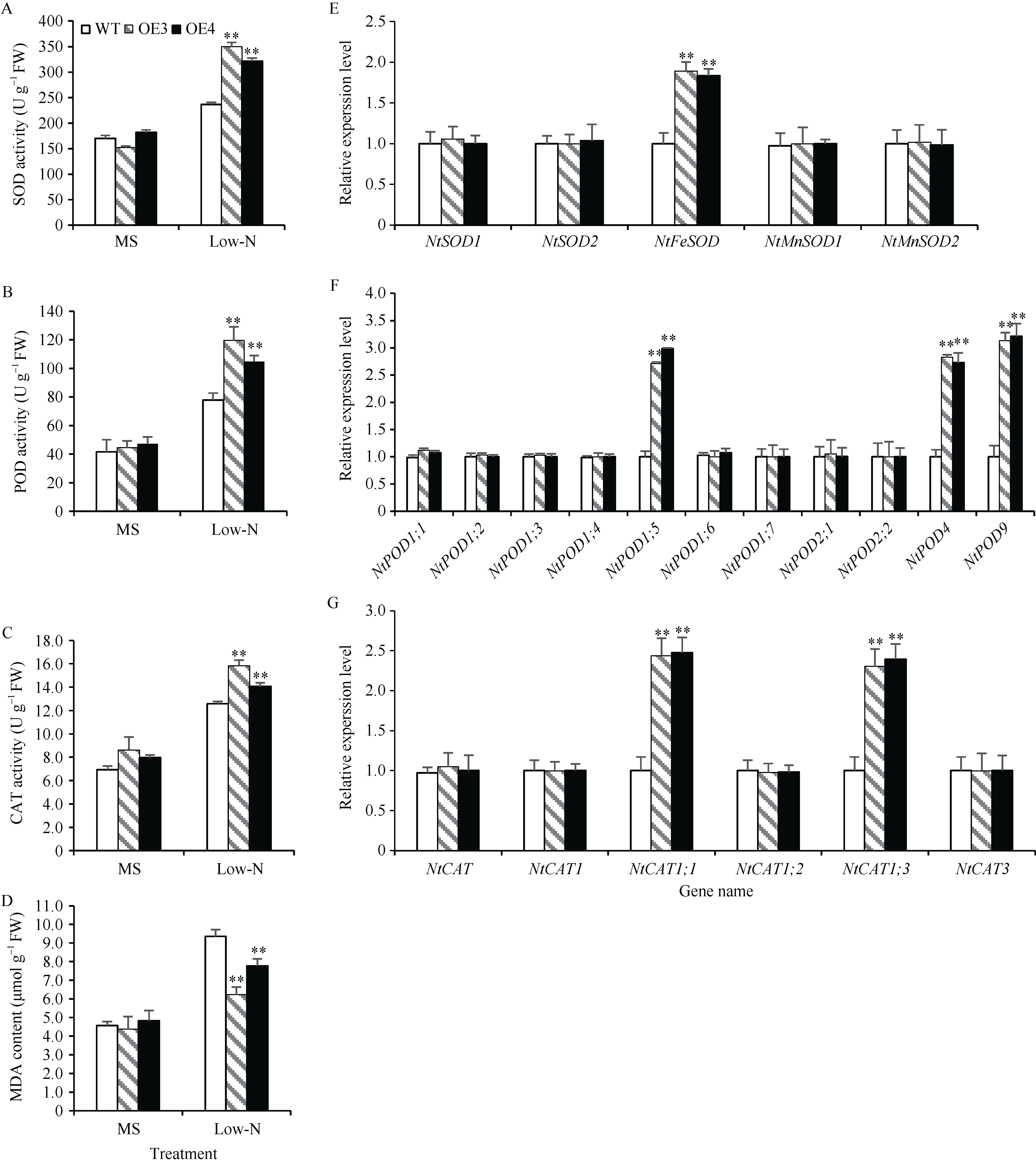
包含SPX、SPX-EXS、SPX-MFS和SPX-RING四个亚族的植物SPX基因家族在磷信号应答中发挥重要功能, 但迄今对小麦的该家族基因成员功能了解尚少。本研究前期从小麦(Triticum aestivum)中鉴定得到一个SPX亚族成员基因TaSPX1 (GenBank No. Ak332300), 亚细胞定位分析发现其定位于细胞核。对TaSPX1和来自小麦、拟南芥和水稻SPX家族的同源蛋白进行系统进化分析, 结果表明, 其与水稻SPX亚族的OsSPX1亲缘关系较近。应用RT-qPCR技术研究发现, TaSPX1的表达量在低氮胁迫下显著增加。构建烟草(Nicotiana tabacum)过表达转基因系(overexpression lines, OE), 应用MS营养液培养对野生型(WT)和OE株系OE3和OE4植株表型进行鉴定。发现在低氮胁迫下, OE3和OE4较WT表现明显的生长优势, 植株鲜重、根重和叶面积显著增加; 包括光合速率、胞间CO2浓度、气孔导度和蒸腾速率在内的光合参数, 以及氮含量、可溶性糖、可溶性蛋白和叶绿素含量也都较WT显著增加。对氮吸收和同化相关基因的表达和酶活性测定结果表明, 上述基因的部分成员在OE植株中的表达量和氮同化酶活性升高。此外, 对包括SOD、POD和CAT在内的植株活性氧清除相关酶活力和MDA含量测定表明, 与WT相比, OE植株中保护酶的活性均明显提高, MDA含量降低。同时发现OE植株中部分保护酶基因成员的表达水平也较WT明显升高。这些结果初步证实了TaSPX1通过改善光合参数、增强氮吸收和转运以及加强保护酶系统等在介导植株抵御低氮胁迫中发挥重要作用。本项研究对小麦SPX家族成员抵御非生物逆境功能增加了新认识, 为作物抗低氮营养逆境的遗传改良提供了理论依据。
| [1] | Anas M, Liao F, Verma K K, Sarwar M A, Mahmood A, Chen Z L, Li Q, Zeng X P, Liu Y, Li Y R. Fate of nitrogen in agriculture and environment: agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol Res, 2021, 53: 4701-4720. |
| [2] | 宋毅, 李静, 谷贺贺, 陆志峰, 廖世鹏, 李小坤, 丛日环, 任涛, 鲁剑巍. 氮肥用量对冬油菜籽粒产量和品质的影响. 作物学报, 2023, 49: 2002-2011. |
| Song Y, Li J, Gu H H, Lu Z F, Liao S P, Li X K, Cong R H, Ren T, Lu J W. Effects of application of nitrogen on seed yield and quality of winter oilseed rape (Brassica napus L.). Acta Agron Sin, 2023, 49: 2002-2011 (in Chinese with English abstract). | |
| [3] |
Li Y, Zou N, Liang X, Zhou X, Guo S, Wang Y, Qin X, Tian Y, Lin J. Effects of nitrogen input on soil bacterial community structure and soil nitrogen cycling in the rhizosphere soil of Lycium barbarum L. Front Microbiol, 2023, 13: 1070817.
doi: 10.3389/fmicb.2022.1070817 |
| [4] |
Masclaux-Daubresse C, Daniel-Vedele F, Dechorgnat J, Chardon F, Gaufichon L, Suzuki A. Nitrogen uptake, assimilation and remobilization in plants: challenges for sustainable and productive agriculture. Ann Bot, 2010, 105: 1141-1157.
doi: 10.1093/aob/mcq028 |
| [5] |
Wang Y Y, Cheng Y H, Chen K E, Tsay Y F. Nitrate transport, signaling, and use efficiency. Annu Rev Plant Biol, 2018, 69: 85-122.
doi: 10.1146/arplant.2018.69.issue-1 |
| [6] |
Fan X, Naz M, Fan X, Xuan W, Miller A J, Xu G. Plant nitrate transporters: from gene function to application. J Exp Bot, 2017, 68: 2463-2475.
doi: 10.1093/jxb/erx011 pmid: 28158856 |
| [7] | Epstein E. Mineral nutrition of plants: principles and perspectives. For Sci, 1972, 19: 3-10. |
| [8] | Fang X Z, Fang S Q, Ye Z Q, Liu D, Zhao K L, Jin C W. NRT1.1 dual-affinity nitrate transport signaling and its roles in plant abiotic stress resistance. Front Plant Sci, 2021, 12: 71569401-71569412. |
| [9] |
Fan X, Feng H, Tan Y, Xu Y, Miao Q, Xu G. A putative 6-transmembrane nitrate transporter OsNRT1.1b plays a key role in rice under low nitrogen. J Integr Plant Biol, 2016, 58: 590-599.
doi: 10.1111/jipb.v58.6 |
| [10] |
Feng Z Q, Li T, Wang X, Sun W J, Zhang T T, You C X, Wang X F. Identification and characterization of apple MdNLP7 transcription factor in the nitrate response. Plant Sci, 2022, 316: 111158.
doi: 10.1016/j.plantsci.2021.111158 |
| [11] |
Xu N, Wang R, Zhao L, Zhang C, Li Z, Lei Z, Liu F, Guan P, Chu Z, Crawford N M, Wang Y. The Arabidopsis NRG2 protein mediates nitrate signaling and interacts with and regulates key nitrate regulators. Plant Cell, 2016, 28: 485-504.
doi: 10.1105/tpc.15.00567 |
| [12] | Qi J, Yu L, Ding J, Ji C, Wang S, Wang C, Ding G, Shi L, Xu F, Cai H. Transcription factor OsSNAC1 positively regulates nitrate transporter gene expression in rice. Plant Physiol, 2023, 19: kiad290. |
| [13] | Wei S, Li X, Lu Z, Zhang H, Ye X, Zhou Y, Li J, Yan Y, Pei H, Duan F, Wang D, Chen S, Wang P, Zhang C, Shang L, Zhou Y, Yan P, Zhao M, Huang J, Bock R, Qian Q, Zhou W. A transcriptional regulator that boosts grain yields and shortens the growth duration of rice. Science, 2022, 377: 1-10. |
| [14] |
Zhang D, Yang K, Kan Z, Dang H, Feng S, Yang Y, Li L, Hou N, Xu L, Wang X, Malnoy M, Ma F, Hao Y, Guan Q. The regulatory module MdBT2-MdMYB88/MdMYB124-MdNRTs regulates nitrogen usage in apple. Plant Physiol, 2021, 185: 1924-1942.
doi: 10.1093/plphys/kiaa118 |
| [15] | Jiang X, Cui H, Wang Z, Kang J, Yang Q, Guo C. Genome-wide analysis of the lateral organ boundaries domain (LBD) members in alfalfa and the involvement of MsLBD48 in nitrogen assimilation. Int J Mol Sci, 2023, 24: 464402-464416. |
| [16] |
Huang W, Ma D, Xia L, Zhang E, Wang P, Wang M, Guo F, Wang Y, Ni D, Zhao H. Overexpression of CsATG3a improves tolerance to nitrogen deficiency and increases nitrogen use efficiency in Arabidopsis. Plant Physiol Biochem, 2023, 196: 328-338.
doi: 10.1016/j.plaphy.2023.01.057 |
| [17] | Kishorekumar R, Bulle M, Wany A, Gupta K J. An overview of important enzymes involved in nitrogen assimilation of plants. Methods Mol Biol, 2020, 57: 1-13. |
| [18] |
Gao Y, Quan S, Lyu B, Tian T, Liu Z, Nie Z, Qi S, Jia J, Shu J, Groot E, Wu J, Wang Y. Barley transcription factor HvNLP2 mediates nitrate signaling and affects nitrogen use efficiency. J Exp Bot, 2022, 73: 770-783.
doi: 10.1093/jxb/erab245 |
| [19] | Cao J, Zheng X, Xie D, Zhou H, Shao S, Zhou J. Autophagic pathway contributes to low-nitrogen tolerance by optimizing nitrogen uptake and utilization in tomato. Hortic Res, 2022, 9: 1-16. |
| [20] |
Zhang Y, He Z, Qi X, Li M, Liu J, Le S, Chen K, Wang C, Zhou Y, Xu Z, Chen J, Guo C, Tang W, Ma Y, Chen M. Overexpression of MYB-like transcription factor SiMYB30 from foxtail millet (Setaria italica L.) confers tolerance to low nitrogen stress in transgenic rice. Plant Physiol Biochem, 2023, 196: 731-738.
doi: 10.1016/j.plaphy.2023.02.025 |
| [21] | Gao S, Yang Y, Yang Y, Zhang X, Su Y, Guo J, Que Y, Xu L. Identification of low-nitrogen-related miRNAs and their target genes in sugarcane and the role of miR156 in nitrogen assimilation. Int J Mol Sci, 2022, 23: 1318701-1318716. |
| [22] |
Secco D, Wang C, Arpat B A, Wang Z, Poirier Y, Tyerman S D, Wu P, Shou H, Whelan J. The emerging importance of the SPX domain-containing proteins in phosphate homeostasis. New Phytol, 2012, 193: 842-851.
pmid: 22403821 |
| [23] |
Stefanovic A, Ribot C, Rouached H, Wang Y, Chong J, Belbahri L, Delessert S, Poirier Y. Members of the PHO1 gene family show limited functional redundancy in phosphate transfer to the shoot, and are regulated by phosphate deficiency via distinct pathways. Plant J, 2007, 50: 982-994.
pmid: 17461783 |
| [24] |
Zhao P, You Q, Lei M. A CRISPR/Cas9 deletion into the phosphate transporter SlPHO1;1 reveals its role in phosphate nutrition of tomato seedlings. Physiol Plant, 2019, 167: 556-563.
doi: 10.1111/ppl.v167.4 |
| [25] |
Liu J, Fu S, Yang L, Luan M, Zhao F, Luan S, Lan W. Vacuolar SPX-MFS transporters are essential for phosphate adaptation in plants. Plant Signal Behav, 2016, 11: e1213474.
doi: 10.1080/15592324.2016.1213474 |
| [26] |
Guo R, Zhang Q, Ying Y, Liao W, Liu Y, Whelan J, Chuanzao M, Shou H. Functional characterization of the three Oryza sativa SPX-MFS proteins in maintaining phosphate homoeostasis. Plant Cell Environ, 2023, 46: 1264-1277.
doi: 10.1111/pce.14414 |
| [27] |
Kant S, Peng M, Rothstein S J. Genetic regulation by NLA and microRNA827 for maintaining nitrate-dependent phosphate homeostasis in Arabidopsis. PLoS Genet, 2011, 7: e1002021.
doi: 10.1371/journal.pgen.1002021 |
| [28] |
Yue W, Ying Y, Wang C, Zhao Y, Dong C, Whelan J, Shou H. OsNLA1, a RING-type ubiquitin ligase, maintains phosphate homeostasis in Oryza sativa via degradation of phosphate transporters. Plant J, 2017, 90: 1040-1051.
doi: 10.1111/tpj.2017.90.issue-6 |
| [29] |
Puga M I, Mateos I, Charukesi R, Wang Z, Franco-Zorrilla J M, de Lorenzo L, Irigoyen M L, Masiero S, Bustos R, Rodríguez J, Leyva A, Rubio V, Sommer H, Paz-Ares J. SPX1 is a phosphate-dependent inhibitor of phosphate starvation response 1 in Arabidopsis. Proc Natl Acad Sci USA, 2014, 111: 14947-14952.
doi: 10.1073/pnas.1404654111 |
| [30] |
Wang Z, Ruan W, Shi J, Zhang L, Xiang D, Yang C, Li C, Wu Z, Liu Y, Yu Y, Shou H, Mo X, Mao C, Wu P. Rice SPX1 and SPX2 inhibit phosphate starvation responses through interacting with PHR2 in a phosphate-dependent manner. Proc Natl Acad Sci USA, 2014, 111: 14953-14958.
doi: 10.1073/pnas.1404680111 pmid: 25271318 |
| [31] |
Zhao L, Liu F, Xu W, Di C, Zhou S, Xue Y, Yu J, Su Z. Increased expression of OsSPX1 enhances cold/subfreezing tolerance in tobacco and Arabidopsis thaliana. Plant Biotechnol J, 2009, 7: 550-561.
doi: 10.1111/pbi.2009.7.issue-6 |
| [32] |
Wang C, Wei Q, Zhang K, Wang L, Liu F, Zhao L, Tan Y, Di C, Yan H, Yu J, Sun C, Chen W J, Xu W, Su Z. Down-regulation of OsSPX1 causes high sensitivity to cold and oxidative stresses in rice seedlings. PLoS One, 2013, 8: e81849.
doi: 10.1371/journal.pone.0081849 |
| [33] |
Hu B, Jiang Z, Wang W, Qiu Y, Zhang Z, Liu Y, Li A, Gao X, Liu L, Qian Y, Huang X, Yu F, Kang S, Wang Y, Xie J, Cao S, Zhang L, Wang Y, Xie Q, Kopriva S, Chu C. Nitrate-NRT1.1B-SPX4 cascade integrates nitrogen and phosphorus signaling networks in plants. Nat Plants, 2019, 5: 401-413.
doi: 10.1038/s41477-019-0384-1 |
| [34] |
Ueda Y, Kiba T, Yanagisawa S. Nitrate-inducible NIGT1 proteins modulate phosphate uptake and starvation signaling via transcriptional regulation of SPX genes. Plant J, 2020, 102: 448-466.
doi: 10.1111/tpj.v102.3 |
| [35] | Yang J, Zhao X, Chen Y, Li G, Li X, Xia M, Sun Z, Chen Y, Li Y, Yao L, Hou H. Identification, structural, and expression analyses of SPX genes in giant duckweed (Spirodela polyrhiza) reveals its role in response to low phosphorus and nitrogen stresses. Cells, 2022, 11: 116701-116724. |
| [36] | Xing X, Du H, Yang Z, Li X, Kong Y, Li W, Zhang C. GmSPX8, a nodule-localized regulator confers nodule development and nitrogen fixation under phosphorus starvation in soybean. BMC Plant Biol, 2022, 22: 16101-16113. |
| [37] |
Tiwari J K, Buckseth T, Devi S, Varshney S, Sahu S, Patil V U, Zinta R, Ali N, Moudgil V, Singh R K, Rawat S, Dua V K, Kumar D, Kumar M, Chakrabarti S K, Rao A R, Rai A. Physiological and genome-wide RNA-sequencing analyses identify candidate genes in a nitrogen-use efficient potato cv. Kufri Gaurav. Plant Physiol Biochem, 2020, 154: 171-183.
doi: 10.1016/j.plaphy.2020.05.041 |
| [38] | 尚文静, 贾利华, 史磊, 林德立, 刘娜, 郑文明. 小麦低磷响应基因的筛选与表达分析. 中国农业大学学报, 2016, 21(10): 1-10. |
| Shang W J, Jia L H, Shi L, Lin D L, Liu N, Zheng W M. Screening and expression analysis of genes responded to low phosphate in wheat root. J China Agric Univ, 2016, 21(10): 1-10 (in Chinese with English abstract). | |
| [39] |
Kumar A, Sharma M, Gahlaut V, Nagaraju M, Chaudhary S, Kumar A, Tyagi P, Gajula M N V P, Singh K P. Genome-wide identification, characterization, and expression profiling of SPX gene family in wheat. Int J Biol Macromol, 2019, 140: 17-32.
doi: S0141-8130(19)34368-5 pmid: 31419556 |
| [40] | Zhao J, Zhao L Q, Gong X D, Feng S Z, Liu X C. Identifification of homeobox transcription factor family in genome-wide and expression pattern analysis of the members in Setosphaeria turcica. Sci Agric Sin, 2017, 50: 669-678. |
| [41] |
Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) method. Methods, 2001, 25: 402-408.
doi: 10.1006/meth.2001.1262 pmid: 11846609 |
| [42] |
Ding W W, Fang W B, Shi S Y, Zhao Y J, Li X J, Xiao K. Wheat WRKY type transcription factor gene TaWRKY1 is essential in mediating drought tolerance associated with an ABA-dependent pathway. Plant Mol Biol Rep, 2016, 34: 1111-1126.
doi: 10.1007/s11105-016-0991-1 |
| [43] | 蒋明月, 苏晓帅, 张宝华, 李小娟, 肖凯. 小麦TaWRKY46介导转基因烟草耐盐性的功能分析. 农业生物技术学报, 2020, 28: 1733-1746. |
| Jiang M Y, Su X S, Zhang B H, Li X J, Xiao K. Functional analysis of TaWRKY46-mediated salt tolerance in transgenic tobacco. J Agric Biotechnol, 2020, 28: 1733-1746 (in Chinese with English abstract). | |
| [44] |
Stuart N W. Adaptation of the micro-kjeldahl method for the determination of nitrogen in plant tissues. Plant Physiol, 1936, 11: 173-179.
doi: 10.1104/pp.11.1.173 pmid: 16653330 |
| [45] | Park B S, Song J T, Seo H S. Arabidopsis nitrate reductase activity is stimulated by the E3 SUMO ligase AtSIZ1. Nat Commun, 2011, 2: 1-10. |
| [46] |
Ferrari T E, Varner J E. Intact tissue assay for nitrite reductase in barley aleurone layers. Plant Physiol, 1971, 47: 790-794.
doi: 10.1104/pp.47.6.790 pmid: 16657706 |
| [47] | Seabra A R, Silva L S, Carvalho H G. Novel aspects of glutamine synthetase (GS) regulation revealed by a detailed expression analysis of the entire GS gene family of Medicago truncatula under different physiological conditions. BMC Plant Biol, 2013, 13: 13701-13715. |
| [48] |
Huang X S, Luo T, Fu X Z, Fan Q J, Liu J H. Cloning and molecular characterization of a mitogen-activated protein kinase gene from Poncirus trifoliata whose ectopic expression confers dehydration/drought tolerance in transgenic tobacco. J Exp Bot, 2011, 62: 5191-5206.
doi: 10.1093/jxb/err229 |
| [49] |
Liu H, Yang W, Liu D, Han Y, Zhang A, Li S. Ectopic expression of a grapevine transcription factor VvWRKY11 contributes to osmotic stress tolerance in Arabidopsis. Mol Biol Rep, 2011, 38: 417-427.
doi: 10.1007/s11033-010-0124-0 |
| [50] | Xu Z, Raza Q, Xu L, He X, Huang Y, Yi J, Zhang D, Shao H B, Ma H, Ali Z. GmWRKY49, a salt-responsive nuclear protein, improved root length and governed better salinity tolerance in transgenic Arabidopsis. Front Plant Sci, 2018, 9: 80901-80911. |
| [51] |
Quan X, Qian Q, Ye Z, Zeng J, Han Z, Zhang G. Metabolic analysis of two contrasting wild barley genotypes grown hydroponically reveals adaptive strategies in response to low nitrogen stress. J Plant Physiol, 2016, 206: 59-67.
doi: 10.1016/j.jplph.2016.07.020 |
| [1] | 张振, 赵俊晔, 石玉, 张永丽, 于振文. 不同播幅对小麦花后叶片光合特性和产量的影响[J]. 作物学报, 2024, 50(4): 981-990. |
| [2] | 李海芬, 鲁清, 刘浩, 温世杰, 王润风, 黄璐, 陈小平, 洪彦彬. 花生赤霉素3-β-双加氧酶(AhGA3ox)基因家族的全基因组鉴定及表达分析[J]. 作物学报, 2024, 50(4): 932-943. |
| [3] | 许乃银, 金石桥, 晋芳, 刘丽华, 徐剑文, 刘丰泽, 任雪贞, 孙全, 许栩, 庞斌双. 基于SNP标记的小麦品种遗传相似度及其检测准确度分析[J]. 作物学报, 2024, 50(4): 887-896. |
| [4] | 黄宏胜, 张馨月, 居辉, 韩雪. 大气CO2浓度升高背景下冬小麦冠层光谱特征和地上生物量估算[J]. 作物学报, 2024, 50(4): 991-1003. |
| [5] | 王添宁, 冯雅岚, 琚吉浩, 吴毅, 张均, 马超. 小麦及其祖先物种GRF转录因子家族鉴定与表达分析[J]. 作物学报, 2024, 50(4): 897-913. |
| [6] | 齐学礼, 李莹, 李春盈, 韩留鹏, 赵明忠, 张建周. 基于转录组探究外源水杨酸对条锈菌侵染小麦幼苗的缓解效应及差异表达基因分析[J]. 作物学报, 2024, 50(4): 1080-1090. |
| [7] | 郝倩琳, 杨廷志, 吕新茹, 秦慧敏, 王亚林, 贾晨飞, 夏先春, 马武军, 徐登安. 小麦胚芽鞘长度QTL定位和GWAS分析[J]. 作物学报, 2024, 50(3): 590-602. |
| [8] | 赵荣荣, 丛楠, 赵闯. 基于Landsat 8影像提取豫中地区冬小麦和夏玉米分布信息的最佳时相选择[J]. 作物学报, 2024, 50(3): 721-733. |
| [9] | 王瑞, 张福耀, 詹鹏杰, 楚建强, 晋敏姗, 赵威军, 程庆军. 基于RNA-Seq筛选高粱低氮胁迫相关候选基因[J]. 作物学报, 2024, 50(3): 669-685. |
| [10] | 琚吉浩, 马超, 王添宁, 吴毅, 董钟, 方美娥, 陈钰姝, 张均, 付国占. 小麦TaPOD家族的全基因组鉴定及表达分析[J]. 作物学报, 2024, 50(3): 779-792. |
| [11] | 范子培, 李龙, 史雨刚, 孙黛珍, 李超男, 景蕊莲. 小麦TabHLH112-2B基因克隆及每穗小穗数相关功能标记开发[J]. 作物学报, 2024, 50(2): 403-413. |
| [12] | 张康, 聂志刚, 王钧, 李广. 温度升高下APSIM模型春小麦籽粒生长参数敏感性分析及优化[J]. 作物学报, 2024, 50(2): 464-477. |
| [13] | 谭丹, 陈家婷, 郜钰, 张晓军, 李欣, 闫贵云, 李锐, 陈芳, 常利芳, 张树伟, 郭慧娟, 畅志坚, 乔麟轶. 小麦穗型相关生长素通路基因发掘及TaARF23-A与小穗数关联分析[J]. 作物学报, 2024, 50(2): 506-513. |
| [14] | 李艳, 方宇辉, 王永霞, 彭超军, 华夏, 齐学礼, 胡琳, 许为钢. 不同磷胁迫处理转OsPHR2小麦的转录组学分析[J]. 作物学报, 2024, 50(2): 340-353. |
| [15] | 谢炜, 贺鹏, 马宏亮, 雷芳, 黄秀兰, 樊高琼, 杨洪坤. 秋闲期秸秆覆盖与施磷对冬小麦氮素吸收利用的影响[J]. 作物学报, 2024, 50(2): 440-450. |
|