欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (12): 3046-3054.doi: 10.3724/SP.J.1006.2024.41014
孙曼1,2( ), 安朝丹2, 高广奇2, 郭杰1, 杨平2, 蒋枞璁2,*()
), 安朝丹2, 高广奇2, 郭杰1, 杨平2, 蒋枞璁2,*()
SUN Man1,2(), AN Chao-Dan2, GAO Guang-Qi2, GUO Jie1, YANG Ping2, JIANG Cong-Cong2,*()
摘要:
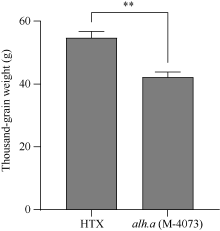
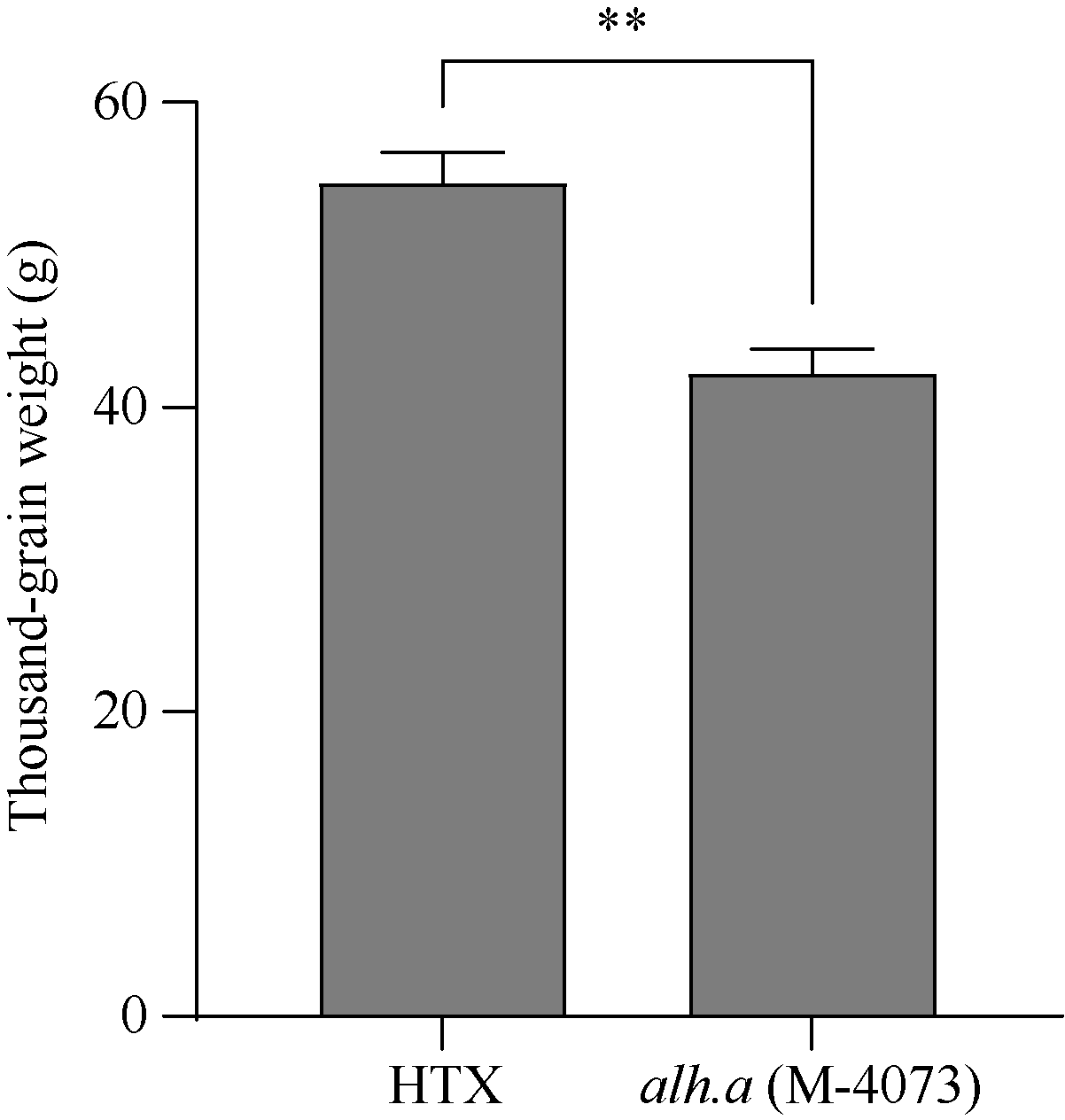
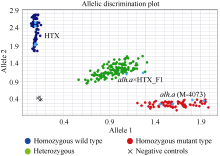
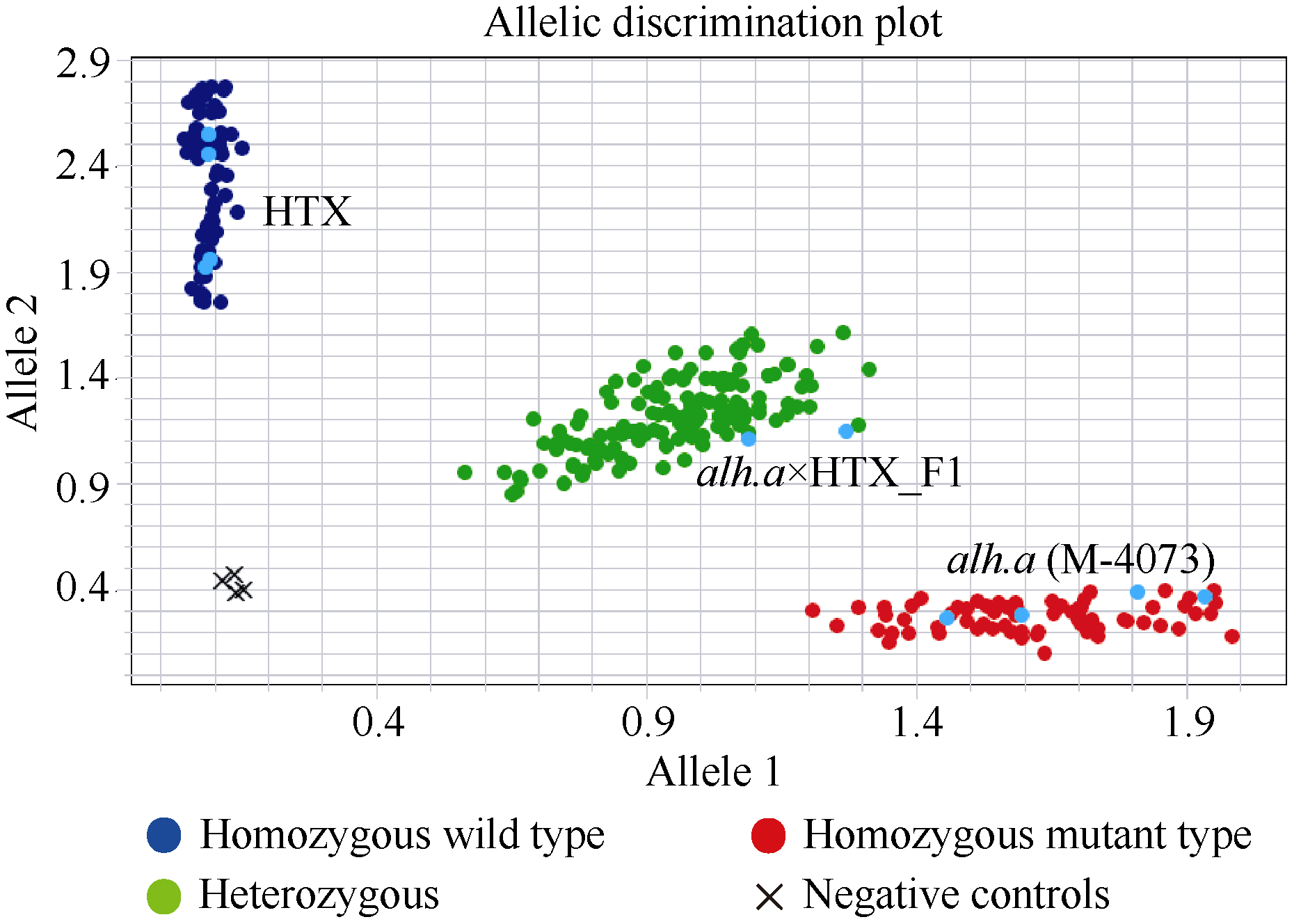
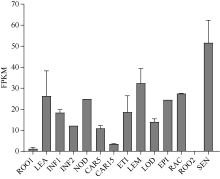
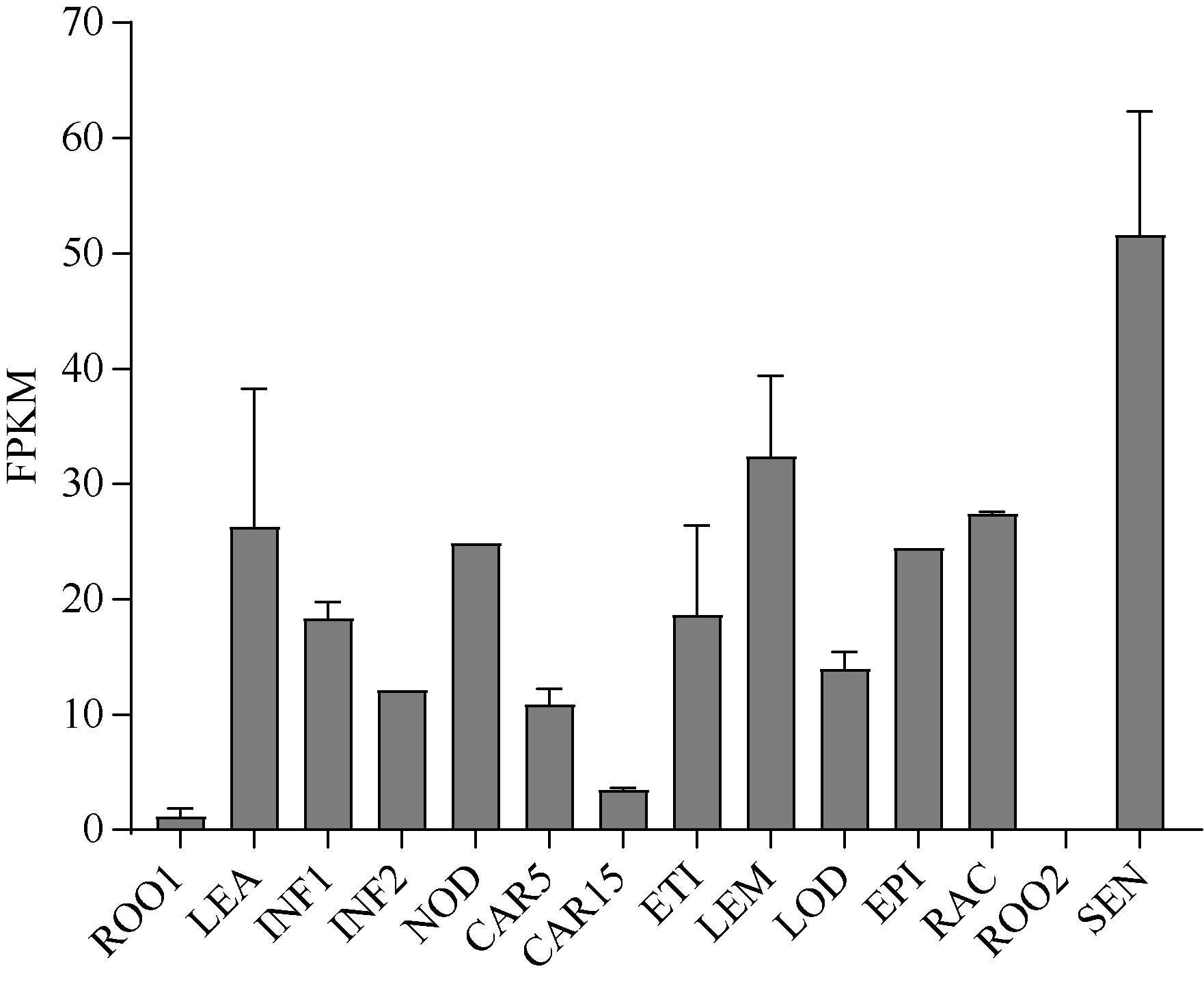
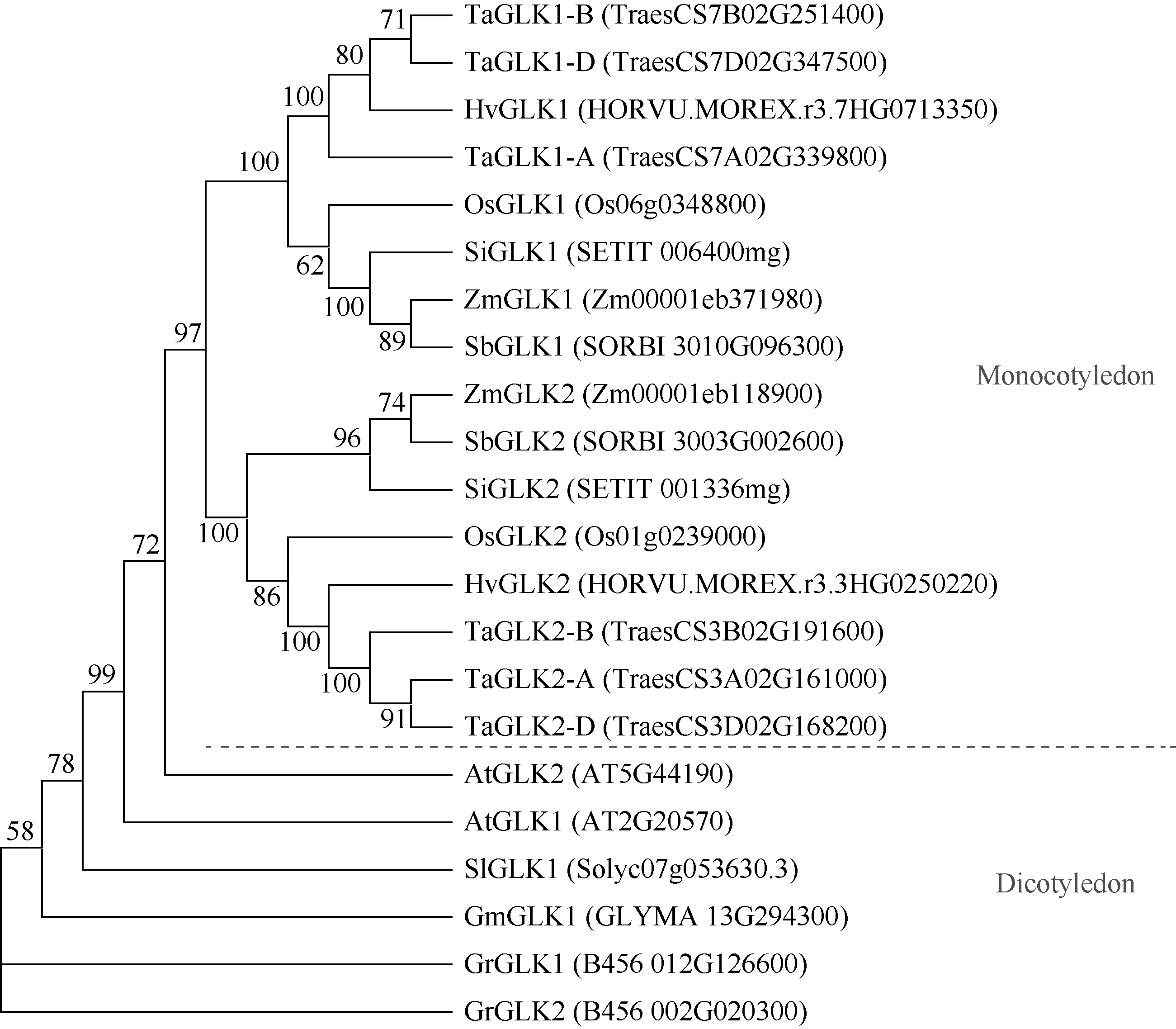
大麦(Hordeum vulgare L.)稃壳是在小穗上包裹种子的花器官, 包括内稃和外稃。大麦稃壳具有光合作用能力, 为籽粒发育提供部分营养物质。通过EMS诱变获得的大麦稃壳白化突变体, 其稃壳明显呈白色, 叶枕、茎节和茎基部均呈白化变异, 但叶片和芒均为绿色。本研究对稃壳白化突变性状进行鉴定和遗传分析, 发现该突变性状由单个隐性基因控制; 通过候选基因测序及竞争性等位基因特异性PCR (KASP)共分离检测, 证实该白化突变是由基因HvGLK2的功能缺失所致; 并在HvGLK2基因上鉴定到3个独立突变事件, 均与已发表的大麦alm1和ebu-a等白化变异不同。HvGLK2基因编码一个Golden 2-like (GLK)转录因子, 为MYB基因家族GARP亚家族成员, 在多数的单、双子叶植物中均存在另一个同源基因GLK1。通过公共数据库检索HvGLK2基因的表达谱数据, 发现HvGLK2在大麦衰老叶片、外稃及穗轴中高丰度表达。本研究明确了HvGLK2基因对大麦稃壳和茎节等组织叶绿素合成的关键作用, 相关的稃壳白化突变体为稃壳光合作用对产量形成的机制研究提供了理想材料。
| [1] | Nishantha M D L C. 野生大麦农艺性状的多样性及其遗传基础. 西北农林科技大学博士学位论文,陕西杨凌, 2018. |
| Nishantha M D L C. Diversity of Wild Barley in Agronomic Traits And Its Genetic Basis. PhD Dissertation of Northwest A&F University, Yangling, Shaanxi, China, 2018 (in Chinese with English abstract). | |
| [2] | Knüpffer H. Genetics and genomics of the triticeae. In: Feuillet C, Muehlbauer G J, eds. Plant Genetics and Genomics: Crops and Models. New York: Springer US, 2009. pp 31-79. |
| [3] | Jiang C C, Lei M M, Guo Y, Gao G Q, Shi L J, Jin Y L, Cai Y, Himmelbach A, Zhou S H, He Q, Yao X F, Kan J H, Haberer G, Duan F Y, Li L H, Liu J, Zhang J, Spannagl M, Liu C M, Stein N, Feng Z Y, Mascher M, Yang P. A reference-guided TILLING by amplicon-sequencing platform supports forward and reverse genetics in barley. Plant Commun, 2022, 3: 100317. |
| [4] | Frey-Wyssling A, Buttrose M S. Photosynthesis in the ear of barley. Nature, 1959, 184: 2031-2032. |
| [5] | Duffus C M, Cochrane M P. Formation of the barley grain- morphology, physiology, and biochemistry. In: MacGregor A W, Bhatty R S, eds. Barley: Chemistry and Technology. St Paul: American Association of Cereal Chemists, 1993. pp 31-72. |
| [6] | Abebe T, Skadsen R W, Kaeppler H F. Cloning and identification of highly expressed genes in barley lemma and Palea. Crop Sci, 2004, 44: 942-950. |
| [7] |
Ziegler-Jöns A. Gas-exchange of ears of cereals in response to carbon dioxide and light: II. Occurrence of a C3-C4 intermediate type of photosynthesis. Planta, 1989, 178: 164-175.
doi: 10.1007/BF00393191 pmid: 24212745 |
| [8] | Wei H, Zhang X Q, Zhu J H, Shang Y, Wang J M, Jia Q J, Zhang Q S, Yang J M, Li C D. Identification and fine mapping of a white husk gene in barley (Hordeum vulgare L.). PLoS One, 2016, 11: e0152128. |
| [9] | Takahashi R, Hayashi J. Linkage study of albino lemma character in barley. Berichte des Ohara Instituts für landwirtschaftliche Biologie, Okayama Universität, 1959, 11: 132-140. |
| [10] |
Taketa S, Hattori M, Takami T, Himi E, Sakamoto W. Mutations in a Golden2-Like gene cause reduced seed weight in barley albino lemma 1 mutants. Plant Cell Physiol, 2021, 62: 447-457.
doi: 10.1093/pcp/pcab001 pmid: 33439257 |
| [11] | Yeh S Y, Lin H H, Chang Y M, Chang Y L, Chang C K, Huang Y C, Ho Y W, Lin C Y, Zheng J Z, Jane W N, Ng C Y, Lu M Y, Lai I L, To K Y, Li W H, Ku M S B. Maize Golden2-like transcription factors boost rice chloroplast development, photosynthesis, and grain yield. Plant Physiol, 2022, 188: 442-459. |
| [12] | 卫云丰. 拟南芥PGA37转录组因子调控叶绿体发育的分子机制. 山西农业大学硕士学位论文,山西太谷, 2020. |
| Wei Y F. Molecular Mechanisms of Arabidopsis PGA37 Transcription Factor in Regulating Chloroplast Development. MS Thesis of Shanxi Agricultural University, Taigu, Shanxi, China, 2020 (in Chinese with English abstract). | |
| [13] | Song J, Wei X J, Shao G N, Sheng Z H, Chen D B, Liu C L, Jiao G A, Xie L H, Tang S Q, Hu P S. The rice nuclear gene WLP1 encoding a chloroplast ribosome L13 protein is needed for chloroplast development in rice grown under low temperature conditions. Plant Mol Biol, 2014, 84: 301-314. |
| [14] | Tsuchiya T. Linkage maps of barley. Barley Genet Newslett, 1980, 10: 95-98. |
| [15] |
金婷, 杨建明, 贾巧君, 汪军妹, 吴宽然, 陈和, 乔海龙, 华为. 大麦白化颖壳突变体的遗传、生理及品质分析. 核农学报, 2013, 27: 1624-1629.
doi: 10.11869/hnxb.2013.11.1624 |
| Jin T, Yang J M, Jia Q J, Wang J M, Wu K R, Chen H, Qiao H L, Hua W. Heredity, physiology and malt quality analysis of albino-lemma barley. J Nucl Agri Sci, 2013, 27: 1624-1629 (in Chinese with English abstract). | |
| [16] | Qin D D, Dong J, Xu F C, Guo G G, Ge S T, Xu Q, Xu Y X, Li M F. Characterization and fine mapping of a novel barley Stage Green-Revertible Albino gene (HvSGRA) by bulked segregant analysis based on SSR assay and specific length amplified fragment sequencing. BMC Genom, 2015, 16: 838. |
| [17] | Faris D G. Physiology and Genetics of the Kernel Color of Barley. PhD Dissertation of University of British Columbia, New York, America, 1955. |
| [18] | Lundqvist U, Franckowiak J D, Konishi T. New and revised descriptions of barley genes. Barley Genet Newslett, 1997, 26: 209. |
| [19] | Yang P, Perovic D, Habekuß A, Zhou R N, Graner A, Ordon F, Stein N. Gene-based high-density mapping of the gene rym7 conferring resistance to Barley mild mosaic virus (BaMMV). Mol Breed, 2013, 32: 27-37. |
| [20] |
安朝丹, 高广奇, 杨平, 程小毛, 蒋枞璁. 大麦钩芒突变体的遗传解析. 植物遗传资源学报, 2023, 24: 1725-1735.
doi: 10.13430/j.cnki.jpgr.20230608001 |
| An C D, Gao G Q, Yang P, Cheng X M, Jiang C C. Genetic dissection of hooded awn mutation in cultivated barley (Hordeum vulgare L.). J Plant Genet Resour, 2023, 24: 1725-1735 (in Chinese with English abstract). | |
| [21] | 高广奇. 大麦黄花叶病抗性新位点rym7H-1遗传解析. 中国农业科学院硕士学位论文,北京, 2021. |
| Gao G Q. Genetic Dissertation of rym7H-1, a New Resistant Locus against BaYMV/BaMMV in Cultivated Barley. MS Thesis of Chinese Academy of Agricultural Sciences,Beijing, China, 2021 (in Chinese with English abstract). | |
| [22] |
Riechmann J L, Heard J, Martin G. Transcription factors: genome- wide comparative analysis among eukaryotes. Science, 2000, 290: 2105-2110.
doi: 10.1126/science.290.5499.2105 pmid: 11118137 |
| [23] | 刘俊芳, 张佳, 李贺, 赵婷婷, 李景富. 植物GOLDEN2-Like转录因子研究进展. 分子植物育种, 2017, 15: 3949-3956. |
| Liu J F, Zhang J, Li H, Zhao T T, Li J F. Research progress of plant GOLDEN2-Like transcription factor. Mol Plant Breed, 2017, 15: 3949-3956 (in Chinese with English abstract). | |
| [24] |
Fitter D W, Martin D J, Copley M J, Scotland R W, Langdale J A. GLK gene pairs regulate chloroplast development in diverse plant species. Plant J, 2002, 31: 713-727.
doi: 10.1046/j.1365-313x.2002.01390.x pmid: 12220263 |
| [25] | Brand A, Borovsky Y, Hill T, Rahman K A A, Bellalou A, van Deynze A, Paran I. CaGLK2 regulates natural variation of chlorophyll content and fruit color in pepper fruit. Theor Appl Genet, 2014, 127: 2139-2148. |
| [26] |
Powell A L T, Nguyen C V, Hill T, Cheng K L, Figueroa-Balderas R, Aktas H, Ashrafi H, Pons C, Fernández-Muñoz R, Vicente A, Lopez-Baltazar J, Barry C S, Liu Y S, Chetelat R, Granell A, van Deynze A, Giovannoni J J, Bennett A B. Uniform ripening encodes a golden 2-like transcription factor regulating tomato fruit chloroplast development. Science, 2012, 336: 1711-1715.
doi: 10.1126/science.1222218 pmid: 22745430 |
| [27] | 刘星, 苏良辰, 张拜宏, 曾丽丹, 李媚娟, 李玲. 异源表达花生基因AhGLK1对拟南芥glk1glk2突变体表型特征及抗旱性的影响. 华南师范大学学报(自然科学版), 2020, 52: 78-84. |
| Liu X, Su L C, Zhang B H, Zeng L D, Li M J, Li L. The effect of heterologous expression of peanut gene AhGLK1 on the phenotypic characteristics and drought resistance of Arabidopsis glk1glk2 mutants. J South China Norm Univ (Nat Sci Edn), 2020, 52: 78-84 (in Chinese with English abstract). |
| [1] | 马敏虎, 常华瑜, 陈朝燕, 仁增, 刘廷辉, 邢国芳, 郭刚刚. 苗草专用型大麦品种鉴定及全基因组关联分析[J]. 作物学报, 2025, 51(1): 91-102. |
| [2] | 鲁宗辉, 司二静, 叶霈颖, 汪军成, 姚立蓉, 马小乐, 李葆春, 王化俊, 尚勋武, 孟亚雄. 大麦籽粒β-葡聚糖含量的全基因组关联分析及候选基因预测[J]. 作物学报, 2024, 50(10): 2483-2492. |
| [3] | 湛潇潇, 冯举伶, 张震欢, 张宏, 汪军成, 李葆春, 杨轲, 司二静, 孟亚雄, 马小乐, 王化俊, 姚立蓉. 大麦HvMBF1c耐盐机制分析[J]. 作物学报, 2024, 50(10): 2503-2514. |
| [4] | 田敏, 刘新春, 潘佳佳, 梁丽静, 董雷, 刘美池, 冯宗云. 大麦籽粒纤维素、半纤维素含量全基因组关联分析[J]. 作物学报, 2023, 49(6): 1726-1732. |
| [5] | 李赢, 刘海翠, 石吕, 石晓旭, 韩笑, 刘建, 魏亚凤. 江苏裸大麦种质资源遗传多样性和群体结构分析[J]. 作物学报, 2023, 49(10): 2687-2697. |
| [6] | 郭楠楠, 刘天策, 史硕, 胡心亭, 牛亚丹, 李亮. 长链非编码RNA (LncRNA)在印度梨形孢促进大麦根部生长发育中的调控作用[J]. 作物学报, 2022, 48(7): 1625-1634. |
| [7] | 郭宝健, 王爽, 吕超, 王菲菲, 许如根. HvLBD19基因对大麦不定根发育的调控[J]. 作物学报, 2022, 48(10): 2435-2442. |
| [8] | 贺军与, 钟伟, 陈云琼, 王卫斌, 熊静蕾, 蒋亚丽, 施辉蒙, 陈升位. 大麦籽粒发育进程中7种黄酮类化合物的积累特性分析[J]. 作物学报, 2021, 47(8): 1624-1630. |
| [9] | 耿腊, 黄业昌, 李梦迪, 谢尚耿, 叶玲珍, 张国平. 大麦籽粒β-葡聚糖含量的全基因组关联分析[J]. 作物学报, 2021, 47(7): 1205-1214. |
| [10] | 张帆, 杨茜. 大麦-双季稻轮作体系有机物料与化肥配施对大麦资源利用效率及产量的影响[J]. 作物学报, 2021, 47(12): 2522-2531. |
| [11] | 徐婷婷, 汪巧玲, 邹淑琼, 狄佳春, 杨欣, 朱银, 赵涵, 颜伟. 基于高通量测序的大麦InDel标记开发及应用[J]. 作物学报, 2020, 46(9): 1340-1350. |
| [12] | 徐银萍, 潘永东, 刘强德, 姚元虎, 贾延春, 任诚, 火克仓, 陈文庆, 赵锋, 包奇军, 张华瑜. 大麦种质资源成株期抗旱性鉴定及抗旱指标筛选[J]. 作物学报, 2020, 46(3): 448-461. |
| [13] | 杨晓梦, 李霞, 普晓英, 杜娟, Muhammad Kazim Ali, 杨加珍, 曾亚文, 杨涛. 大麦重组自交系群体籽粒总花色苷含量和千粒重QTL定位[J]. 作物学报, 2020, 46(01): 52-61. |
| [14] | 时丽洁,蒋枞璁,王方梅,杨平,冯宗云. 大麦蛋白质二硫键异构酶基因家族的鉴定与表达分析[J]. 作物学报, 2019, 45(9): 1365-1374. |
| [15] | 张莉莎,米胜南,王玲,委刚,郑尧杰,周恺,尚丽娜,朱美丹,王楠. 水稻短根白化突变体sra1生理生化分析及基因定位[J]. 作物学报, 2019, 45(4): 556-567. |
|