欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (5): 807-813.doi: 10.3724/SP.J.1006.2021.03039
葛敏( ), 王元琮, 宁丽华, 胡梦梅, 石习, 赵涵*()
), 王元琮, 宁丽华, 胡梦梅, 石习, 赵涵*()
GE Min(), WANG Yuan-Cong, NING Li-Hua, HU Meng-Mei, SHI Xi, ZHAO Han*()
摘要:
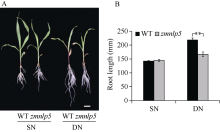
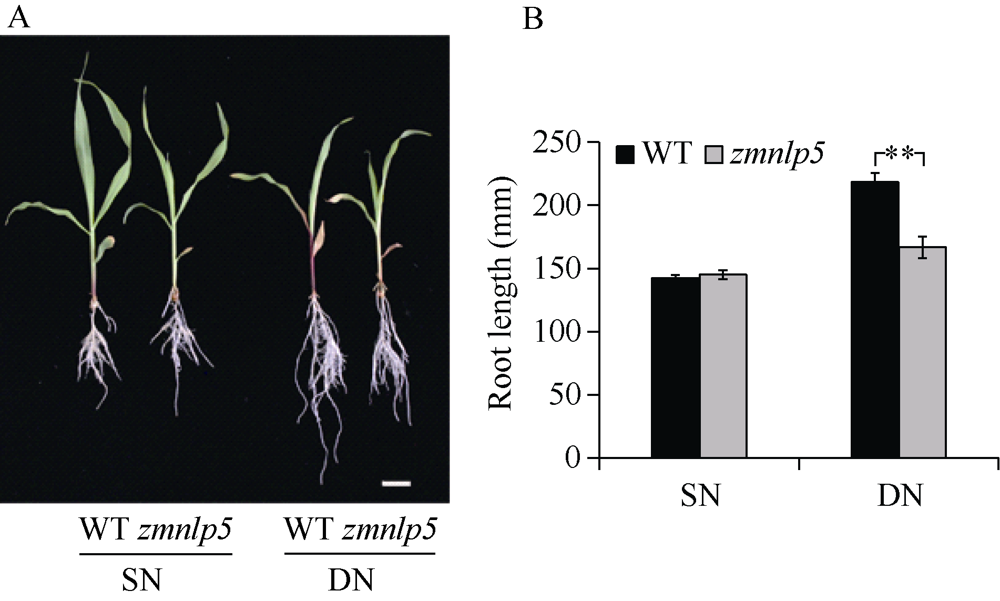
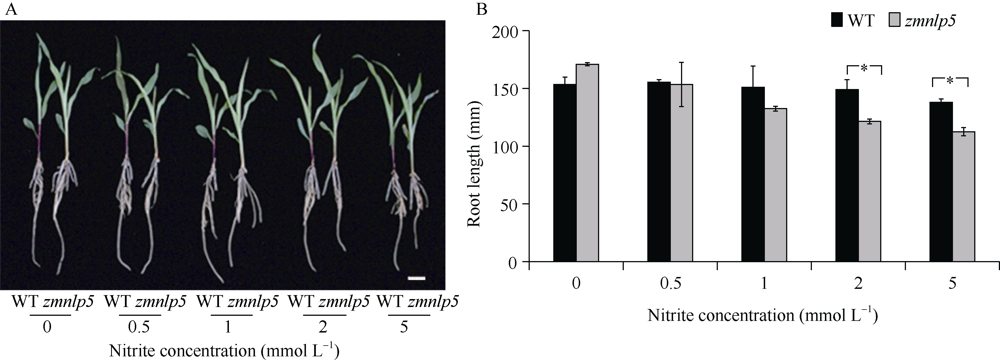
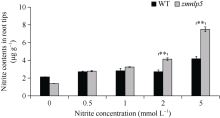
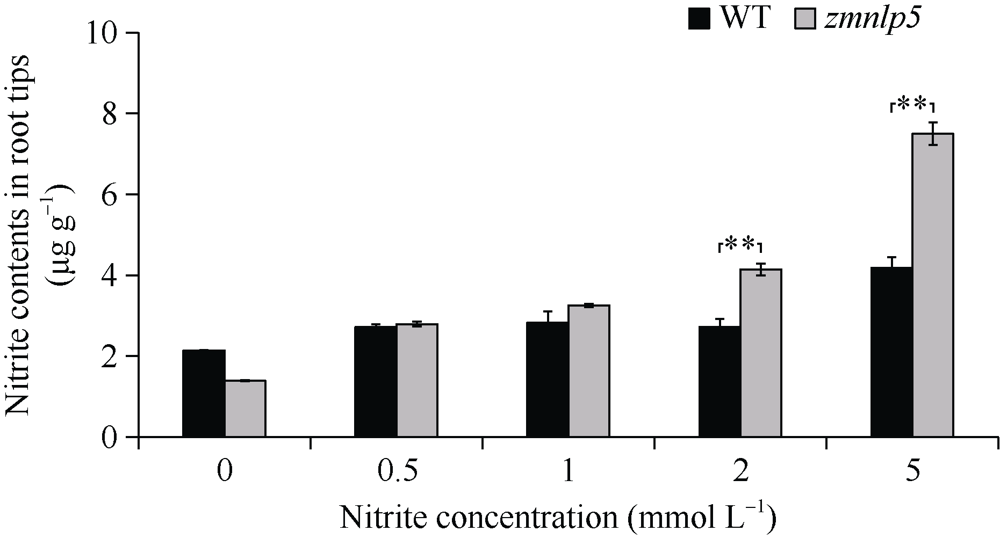
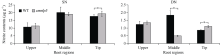
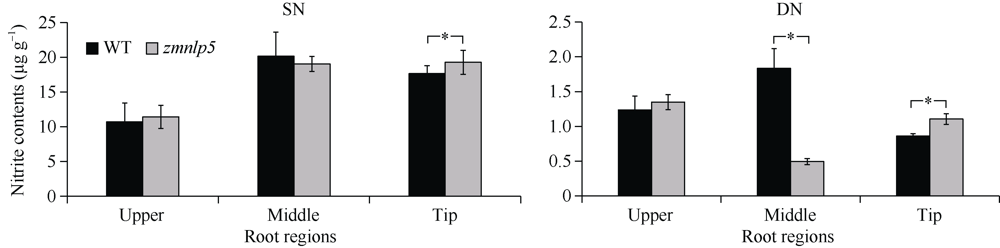
研究作物氮高效机制并挖掘利用其中重要的调控基因, 对我国农业绿色发展具有重要意义。前期我们发现氮响应转录因子ZmNLP5调控亚硝酸还原酶基因的表达, 对玉米氮素吸收利用具有促进作用, 但其调控机制尚未明确。基于此, 本研究以ZmNLP5基因突变体(zmnlp5)和野生型(WT)植株为研究材料, 深入解析ZmNLP5影响玉米氮素吸收利用的生理机制。足氮(sufficient nitrogen, SN)和低氮(deficient nitrogen, DN)条件下研究材料表型分析发现, DN条件下相对于WT植株, zmnlp5植株根长显著降低。根部不同区域ZmNLP5表达量分析发现, ZmNLP5主要在根尖区域表达。不同亚硝酸盐浓度处理下, 研究材料的根长和根尖区域亚硝酸盐含量分析发现, 当亚硝酸盐的浓度高于2 mmol L-1时, zmnlp5突变体根长相对于WT显著降低, zmnlp5植株根尖区域亚硝酸盐积累量显著高于WT。SN和DN条件下研究材料根部亚硝酸盐含量检测发现, DN条件zmnlp5植株根尖区域所积累的亚硝酸盐也显著高于WT。综上所述, 转录因子ZmNLP5在玉米植株响应低氮环境根部伸长生长中发挥重要功能, 该结果为玉米氮高效育种提供了一定的理论基础。
| [1] | Gutierrez R A. Systems biology for enhanced plant nitrogen nutrition. Science, 2012,336:1673-1675. |
| [2] | Guana P Z, Ripolla J J, Wang R H, Vuong L, Bailey-Steinitz L J, Ye D, Crawford N M. Interacting TCP and NLP transcription factors control plant responses to nitrate availability. Proc Natl Acad Sci USA, 2017,114:2419-2424. |
| [3] | Bi Y M, Meyer A, Downs G S, Shi X, El-Kereamy A, Lukens L, Rothstein S J. High throughput RNA sequencing of a hybrid maize and its parents shows different mechanisms responsive to nitrogen limitation. BMC Genom, 2014,15:77. |
| [4] | Humbert S, Subedi S, Cohn J, Zeng B, Bi Y M, Chen X, Zhu T, McNicholas P D, Rothstein S J. Genome-wide expression profiling of maize in response to individual and combined water and nitrogen stresses. BMC Genom, 2013,14:3. |
| [5] | Xu G, Fan X, Miller A J. Plant nitrogen assimilation and use efficiency. Annu Rev Plant Biol, 2012,63:153-182. |
| [6] | Crawford N M. Nitrate: nutrient and signal for plant growth. Plant Cell, 1995,7:859-868. |
| [7] | Crawford N M, Forde B G. Molecular and developmental biology of inorganic nitrogen nutrition. Arabid Book, 2002,1:e0011. |
| [8] | Moose S, Below F E. Biotechnology approaches to improving maize nitrogen use efficiency. In: Kriz A, Larkins B, eds. Molecular Genetic Approaches to Maize Improvement. Biotechnology in Agriculture and Forestry, Springer, Berlin, Heidelberg, 2009. pp 65-77. |
| [9] | Ueda Y, Konishi M, Yanagisawa S. Molecular basis of the nitrogen response in plants. Soil Sci Plant Nutr, 2017,63:329-341. |
| [10] | Yu P, Eggert K, von Wirén N, Li C J, Hochholdinger F. Cell type-specific gene expression analyses by RNA sequencing reveal local high nitrate-triggered lateral root initiation in shoot-borne roots of maize by modulating auxin-related cell cycle regulation. Plant Physiol, 2015,169:690. |
| [11] | Trevisan S, Manoli A, Ravazzolo L, Botton A, Pivato M, Masi A, Quaggiotti S. Nitrate sensing by the maize root apex transition zone: a merged transcriptomic and proteomic survey. J Exp Bot, 2015,66:3699-3715. |
| [12] | Gutierrez R A, Stokes T L, Thum K, Xu X, Obertello M, Katari M S, Tanurdzic M, Dean A, Nero D C, McClung C R, Coruzzi G M. Systems approach identifies an organic nitrogen- responsive gene network that is regulated by the master clock control gene CCA1. Proc Natl Acad Sci USA, 2008,105:4939-4944. |
| [13] | Rubin G, Tohge T, Matsuda F, Saito K, Scheible W R. Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant Cell, 2009,21:3567-3584. |
| [14] | Marchive C, Roudier F, Castaings L, Brehaut V, Blondet E, Colot V, Meyer C, Krapp A. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat Commun, 2013,4:1713. |
| [15] | Ge M, Liu Y H, Jiang L, Wang Y C, Lyu Y D, Zhou L, Liang S Q, Bao H B, Zhao H. Genome-wide analysis of maize NLP transcription factor family revealed the roles in nitrogen response. Plant Growth Regul, 2018,84:95-105. |
| [16] | Konishi M, Yanagisawa S. Identification of a nitrate-responsive cis-element in the Arabidopsis NIR1 promoter defines the presence of multiple cis-regulatory elements for nitrogen response. Plant J, 2010,63:269-282. |
| [17] | Ge M, Wang Y C, Liu Y H, Jiang L, He B, Ning L H, Du H Y, Lyu Y D, Zhou L, Lin F, Zhang T F, Liang S Q, Lu H Y, Zhao H. The NIN-like protein 5 (ZmNLP5) transcription factor is involved in modulating the nitrogen response in maize. Plant J, 2020,102:353-368. |
| [18] | Hachiya T, Ueda N, Kitagawa M, Hanke G, Suzuki A, Hase T, Sakakibara H. Arabidopsis root type ferredoxin: NADP(H) oxidoreductase 2 is involved in detoxification of nitrite in roots. Plant Cell Physiol, 2016,57:2440-2450. |
| [19] | Schlüter U, Mascher M, Colmsee C, Scholz U, Bräutigam A, Fahnenstich H, Sonnewald U. Maize source leaf adaptation to nitrogen deficiency affects not only nitrogen and carbon metabolism but also control of phosphate homeostasis. Plant Physiol, 2012,160:1384-1406. |
| [20] | Lin F, Jiang L, Liu Y, Lv Y, Dai H, Zhao H. Genome-wide identification of housekeeping genes in maize. Plant Mol Biol, 2014,86:543-554. |
| [21] | Zanin L, Zamboni A, Monte R, Tomasi N, Varanini Z, Cesco S, Pinton R. Transcriptomic analysis highlights reciprocal interactions of urea and nitrate for nitrogen acquisition by maize roots. Plant Cell Physiol, 2015,56, 532-548. |
| [22] | Morot-Gaudry-Talarmain Y, Rockel P, Moureaux T, Quilleré I, Leydecker M, Kaiser W, Morot-Gaudry J F. Nitrite accumulation and nitric oxide emission in relation to cellular signaling in nitrite reductase antisense tobacco. Planta, 2002,215:708-715. |
| [23] | Kiba T, Krapp A. Plant nitrogen acquisition under low availability: regulation of uptake and root architecture. Plant Cell Physiol, 2016,57:707-714. |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [4] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [5] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [6] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [7] | 郭星宇, 刘朋召, 王瑞, 王小利, 李军. 旱地冬小麦产量、氮肥利用率及土壤氮素平衡对降水年型与施氮量的响应[J]. 作物学报, 2022, 48(5): 1262-1272. |
| [8] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [9] | 孙思敏, 韩贝, 陈林, 孙伟男, 张献龙, 杨细燕. 棉花苗期根系分型及根系性状的关联分析[J]. 作物学报, 2022, 48(5): 1081-1090. |
| [10] | 朱峥, 王田幸子, 陈悦, 刘玉晴, 燕高伟, 徐珊, 马金姣, 窦世娟, 李莉云, 刘国振. 水稻转录因子WRKY68在Xa21介导的抗白叶枯病反应中发挥正调控作用[J]. 作物学报, 2022, 48(5): 1129-1140. |
| [11] | 陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展[J]. 作物学报, 2022, 48(4): 781-790. |
| [12] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [13] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [14] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [15] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
|
||