欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (10): 2575-2587.doi: 10.3724/SP.J.1006.2022.14174
孙棋棋( ), 郑永美, 于天一, 吴月, 杨吉顺, 吴正锋(), 吴菊香, 李尚霞
), 郑永美, 于天一, 吴月, 杨吉顺, 吴正锋(), 吴菊香, 李尚霞
SUN Qi-Qi(), ZHENG Yong-Mei, YU Tian-Yi, WU Yue, YANG Ji-Shun, WU Zheng-Feng(), WU Ju-Xiang, LI Shang-Xia
摘要:
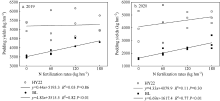
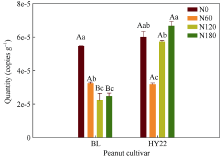
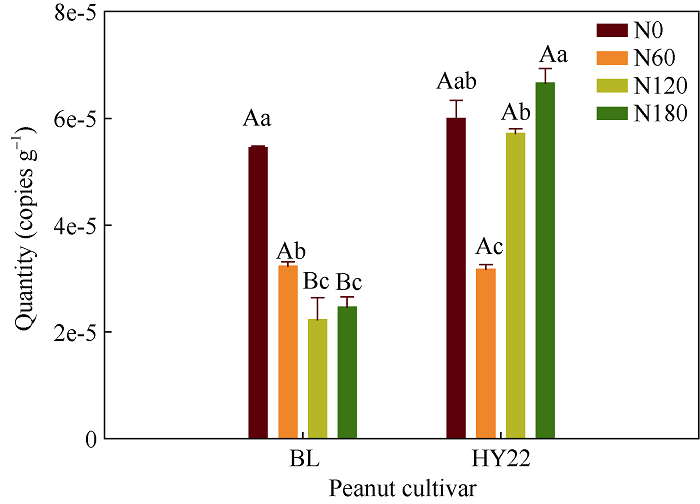
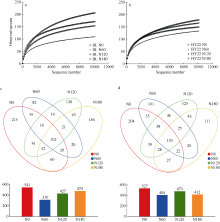
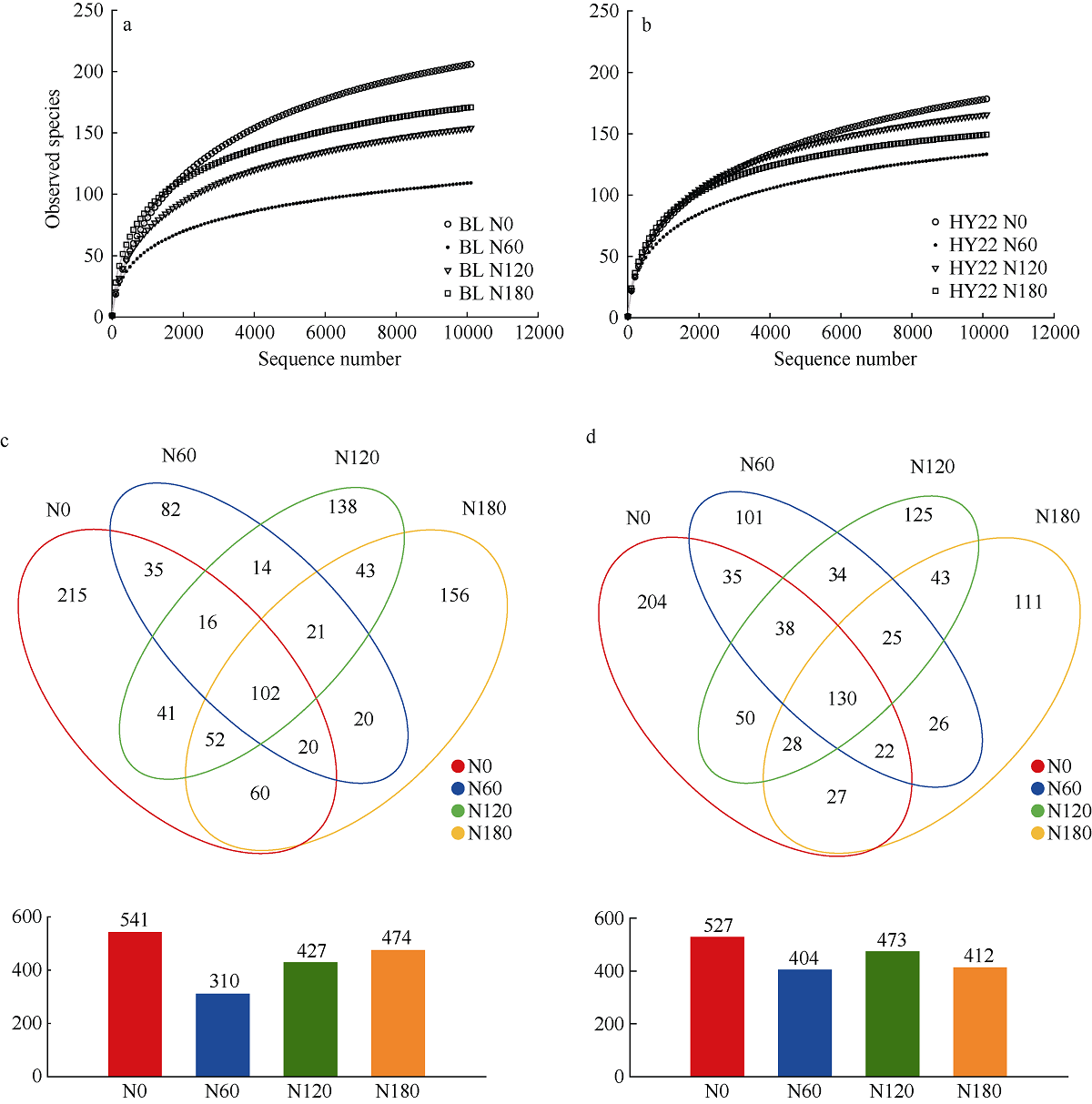
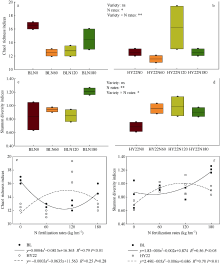
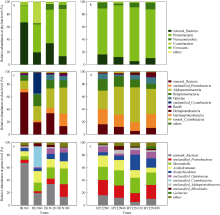
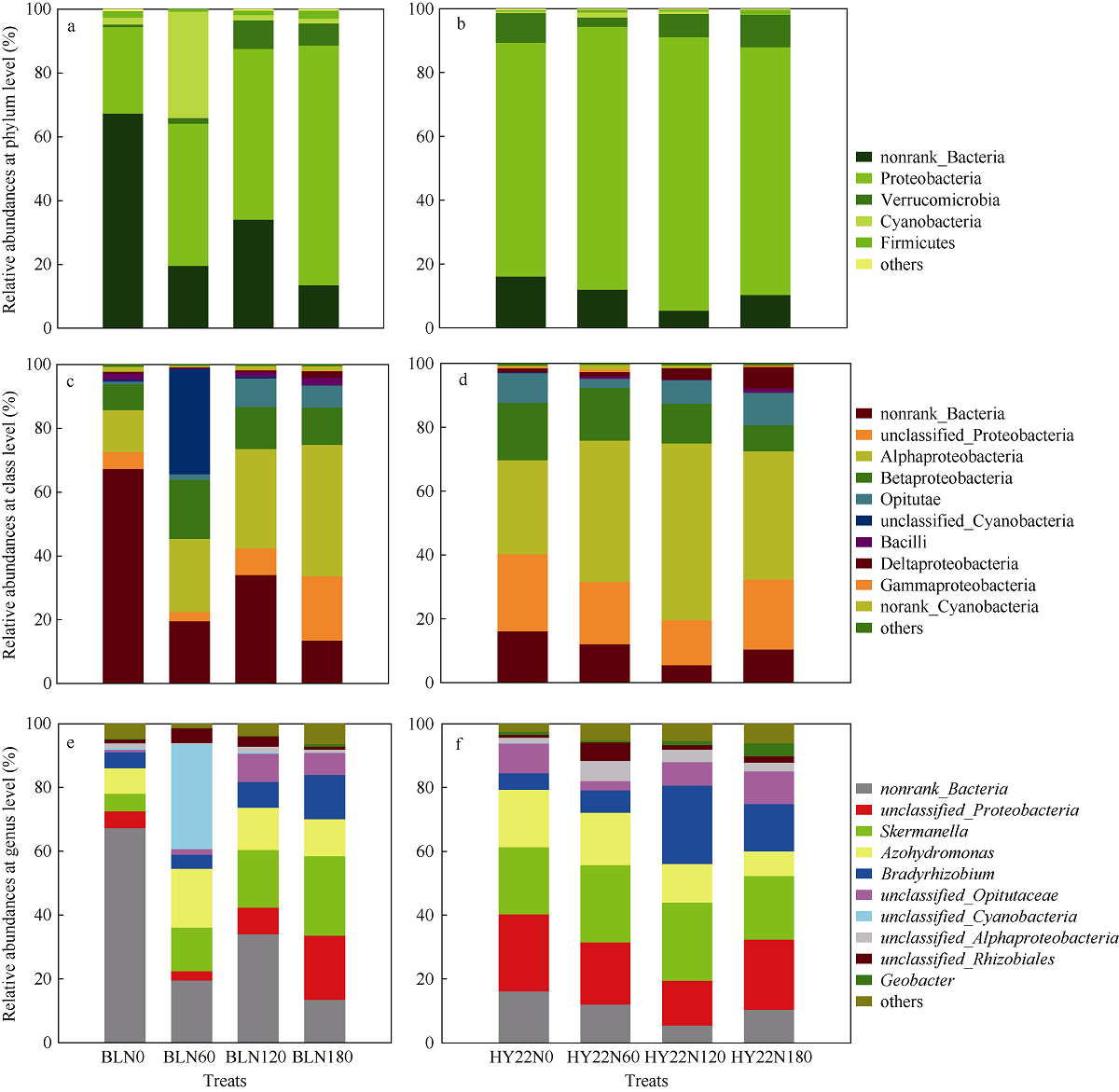
施氮对豆科作物土壤固氮菌的影响机理尚不明确。深入研究施氮对花生土壤固氮菌的影响, 对花生田高效施氮和农业可持续发展具有重要意义。本研究以结瘤(花育22)和不结瘤(BL)花生为材料, 设置N0 (不施氮, 0 kg hm-2)、N60 (减量施氮, 60 kg hm-2)、N120 (常量施氮, 120 kg hm-2)和N180 (过量施氮, 180 kg hm-2) 4个施氮水平, 探讨不同施氮水平对不同结瘤特性花生土壤固氮菌的影响。采用实时荧光定量PCR和高通量测序技术, 以nifH基因为标靶, 分析土壤固氮菌丰度、多样性和群落组成。结果表明: (1) 施氮显著提高土壤氮组分含量, N120结瘤花生土壤中微生物量氮和可溶性有机氮显著高于不结瘤花生。不结瘤花生荚果产量随施氮水平呈线性增加, 而结瘤花生产量不受施氮影响。(2) 施氮条件下, 不结瘤花生土壤nifH拷贝数显著降低, 而结瘤花生土壤nifH拷贝数随施氮水平增加先降低后增加。(3) 施氮对不结瘤花生土壤固氮菌多样性先抑制后促进, 而施氮对结瘤花生土壤固氮菌多样性呈先促进后抑制的模式, 并在N120达到最大值。(4) 施氮显著改变不结瘤花生土壤固氮菌群落组成, 不同施氮水平优势属分别为nonrank_Bacteria (N0)、unclassified_Cyanobacteria (N60)、nonrank_Bacteria (N120)和Skermanella (N180), 硝态氮是影响处理间土壤固氮菌群落组成差异的唯一显著因素; 结瘤花生土壤各施氮水平固氮菌群落组成相似, 除N120外, 均以unclassified_Proteobacteria和Skermanella占优。不结瘤与结瘤花生土壤固氮菌对施氮水平截然不同的响应可能是由于二者分别受到不同氮源的影响(肥料氮vs.根瘤固氮)。本研究中120 kg hm-2是对土壤固氮和花生生产最优的施氮水平。
| [1] |
Cleveland C C, Townsend A R, Schimel D S, Fisher H, Howarth R W, Hedin L O, Perakis S S, Latty E F, Fischer J C V, Elseroad A. Global patterns of terrestrial biological nitrogen (N2) fixation in natural ecosystems. Global Biogeochem Cycles, 1999, 13: 623-645.
doi: 10.1029/1999GB900014 |
| [2] |
Roesch L, Camargo F, Bento F M, Triplett E W. Biodiversity of diazotrophic bacteria within the soil, root and stem of field-grown maize. Plant Soil, 2008, 302: 91-104.
doi: 10.1007/s11104-007-9458-3 |
| [3] |
Hsu S F, Buckley D H. Evidence for the functional significance of diazotroph community structure in soil. ISME J, 2008, 3: 124-136.
doi: 10.1038/ismej.2008.82 |
| [4] |
Li Y, Pan F, Yao H. Response of symbiotic and asymbiotic nitrogen-fixing microorganisms to nitrogen fertilizer application. J Soils Sediments, 2019, 19: 1948-1958.
doi: 10.1007/s11368-018-2192-z |
| [5] | 郑棉海, 陈浩, 朱晓敏, 毛庆功, 莫江明. 矿质养分输入对森林生物固氮的影响. 生态学报, 2015, 35: 7941-7954. |
| Zheng M H, Chen H, Zhu X M, Mao Q G, Mo J M. Effect of mineral nutrient input on biological nitrogen fixation in forest. Acta Ecol Sin, 2015, 35: 7941-7954. (in Chinese with English abstract) | |
| [6] | Chen L, Li K K, Shi W J, Wang X L, Chen W X. Negative impacts of excessive nitrogen fertilization on the abundance and diversity of diazotrophs in black soil under maize monocropping. Geoderma, 2021, 393: 114999. |
| [7] |
Wang C, Zheng M, Song W, Wen S, Wang B, Zhu C, Shen R. Impact of 25 years of inorganic fertilization on diazotrophic abundance and community structure in an acidic soil in southern China. Soil Biol Biochem, 2017, 113: 240-249.
doi: 10.1016/j.soilbio.2017.06.019 |
| [8] |
Reed R, Marjon D V, Portilho C N, Evódio M, Edilson P, Lucy S. Diversity of nifH gene pools in the rhizosphere of two cultivars of sorghum (Sorghum bicolor) treated with contrasting levels of nitrogen fertilizer. FEMS Microbiol Lett, 2010, 279: 15-22.
doi: 10.1111/j.1574-6968.2007.00975.x |
| [9] | Fan K, Delgado-Baquerizo M, Guo X, Wang D, Wu Y, Zhu M, Yu W, Yao H, Zhu Y G, Chu H. Suppressed N fixation and diazotrophs after four decades of fertilization. Microbiome, 2019, 7: 143. |
| [10] |
Wang J, Li Q, Shen C, Yang F, Wang J, Yuan G. Significant dose effects of fertilizers on soil diazotrophic diversity, community composition, and assembly processes in a long-term paddy field fertilization experiment. Land Degrad Develop, 2021, 32: 1-10.
doi: 10.1002/ldr.3658 |
| [11] |
Sun Q, Rui W, Ying W, Du L, Man Z, Xin G, Hu Y, Guo S. Temperature sensitivity of soil respiration to nitrogen and phosphorous fertilization: Does soil initial fertility matter? Geoderma, 2018, 325: 172-182.
doi: 10.1016/j.geoderma.2018.04.001 |
| [12] |
Liao H, Li Y, Yao H. Fertilization with inorganic and organic nutrients changes diazotroph community composition and N-fixation rates. J Soils Sediments, 2017, 18: 1076-1086.
doi: 10.1007/s11368-017-1836-8 |
| [13] | 陈洁, 骆土寿, 周璋, 许涵, 陈德祥, 李意德. 氮沉降对热带亚热带森林土壤氮循环微生物过程的影响研究进展. 生态学报, 2020, 40: 8528-8538. |
| Chen J, Luo T S, Zhou Z, Xu H, Chen D X, Li Y D. Research advances in nitrogen deposition effects on microbial processes involved in soil nitrogen cycling in tropical and subtropical forests. Acta Ecol Sin, 2020, 40: 8528-8538. (in Chinese with English abstract) | |
| [14] |
Jacot K A, Lüscher A, NöSberger J, Hartwig U A. Symbiotic N2 fixation of various legume species along an altitudinal gradient in the Swiss Alps. Soil Biol Biochem, 2000, 32: 1043-1052.
doi: 10.1016/S0038-0717(00)00012-2 |
| [15] |
Rösch C, Mergel A, Hermann Bothe H. Biodiversity of denitrifying and dinitrogen-fixing bacteria in an acid forest soil. Appl Environ Microbiol, 2002, 68: 3818-3829.
doi: 10.1128/AEM.68.8.3818-3829.2002 |
| [16] | Xiao D, Tan Y, Liu X, Yang R, Wang K. Responses of soil diazotrophs to legume species and density in a karst grassland, southwest China. Agric Ecosyst Environ, 2020, 288: 106707. |
| [17] |
Wang C B, Zheng Y M, Shen P, Zheng Y P, Wu Z F, Sun X W, Yu T Y, Feng H. Determining N supplied sources and N use efficiency for peanut under applications of four forms of N fertilizers labeled by isotope 15N. J Integr Agric, 2016, 15: 432-439.
doi: 10.1016/S2095-3119(15)61079-6 |
| [18] | 吴正锋, 陈殿绪, 郑永美, 王才斌, 孙学武, 李向东, 王兴祥, 石程仁, 冯昊. 花生不同氮源供氮特性及氮肥利用率研究. 中国油料作物学报, 2016, 38: 207-213. |
| Wu Z F, Chen D X, Zheng Y M, Wang C B, Sun X W, Li X D, Wang X X, Shi C R, Feng H. Supply characteristics of different nitrogen sources and nitrogen use efficiency of peanut. Chin J Oil Crop Sci, 2016, 38: 207-213. (in Chinese with English abstract) | |
| [19] |
郑永美, 杜连涛, 王春晓, 吴正锋, 孙学武, 于天一, 沈浦, 王才斌. 不同花生品种根瘤固氮特点及其与产量的关系. 应用生态学报, 2019, 30: 961-968.
pmid: 30912389 |
|
Zheng Y M, Du L T, Wang C X, Wu Z F, Sun X W, Yu T Y, Shen P, Wang C B. Nitrogen fixation characteristics of root nodules in different peanut varieties and their relationship with yield. Chin J Appl Ecol, 2019, 30: 961-968. (in Chinese with English abstract)
doi: 10.13287/j.1001-9332.201903.019 pmid: 30912389 |
|
| [20] | 左元梅, 刘永秀, 张福锁. NO3-态氮对花生结瘤与固氮作用的影响. 生态学报, 2003, 23: 758-764. |
| Zuo Y M, Liu Y X, Zhang F S. Effects of the NO3--N on nodule formation and nitrogen fixing of peanut. Acta Ecol Sin, 2003, 23: 758-764. (in Chinese with English abstract) | |
| [21] |
Lin Y, Ye G, Liu D, Ledgard S, Luo J, Fan J, Yuan J, Chen Z, Ding W. Long-term application of lime or pig manure rather than plant residues suppressed diazotroph abundance and diversity and altered community structure in an acidic Ultisol. Soil Biol Biochem, 2018, 123: 218-228.
doi: 10.1016/j.soilbio.2018.05.018 |
| [22] | 吴海宁, 黄志鹏, 唐秀梅, 熊发前, 钟瑞春, 贺梁琼, 韩柱强, 蒋菁, 刘菁, 唐荣华. 氮肥减施对花生根际土壤固氮微生物多样性的影响. 江苏农业科学, 2019, 47(16): 93-97. |
| Wu H N, Huang Z P, Tang X M, Xiong F M, Zhong R C, He L Q, Han Z Q, Jiang J, Liu J, Tang R H. Effect of nitrogen reduced-fertilization on soil diazotrophic diversity in peanut rhizosphere. Jiangsu Agric Sci, 2019, 47(16): 93-97. (in Chinese with English abstract) | |
| [23] |
Gorbet D W, Burton J C. A non-nodulating peanut. Crop Sci, 1979, 19: 727-728.
doi: 10.2135/cropsci1979.0011183X001900050045x |
| [24] | 石海, 苗淑杰, 刘居东, 周克琴. 施氮对结瘤和非结瘤近等位基因大豆生长和固氮性状的影响. 大豆科学, 2012, 31: 961-965. |
| Shi H, Miao S J, Liu J D, Zhou K Q. Effect of nitrogen application on growth and nitrogen fixation in nodulation and non-nodulation soybean isolines. Soybean Sci, 2012, 31: 961-965. (in Chinese with English abstract) | |
| [25] |
Selamat A, Gardner F P. Nitrogen partitioning and redistribution in nonnodulating peanut related to nitrogen stress. Agron J, 1985, 77: 859-862.
doi: 10.2134/agronj1985.00021962007700060009xa |
| [26] |
郑永美, 王春晓, 刘岐茂, 吴正锋, 王才斌, 孙秀山, 郑亚萍. 氮肥对花生根系生长和结瘤能力的调控效应. 核农学报, 2017, 31: 2418-2425.
doi: 10.11869/j.issn.100-8551.2017.12.2418 |
| Zheng Y M, Wang C X, Liu Q M, Wu Z F, Wang C B, Sun X S, Zheng Y P. Effect of nitrogen fertilizer regulation on root growth and nodulating ability of peanut. J Nucl Agric Sci, 2017, 31: 2418-2425. (in Chinese with English abstract) | |
| [27] | Wang Q, Wang J, Li Y, Chen D, Ao J, Zhou W, Shen D, Li Q, Huang Z, Jiang Y. Influence of nitrogen and phosphorus additions on N2-fixation activity, abundance, and composition of diazotrophic communities in a Chinese fir plantation. Sci Total Environ, 2017, 619: 1530-1537. |
| [28] | 徐鹏霞, 韩丽丽, 贺纪正, 罗锋, 张丽梅. 非共生生物固氮微生物分子生态学研究进展. 应用生态学报, 2017, 28: 3440-3450. |
| Xu P X, Han L L, He J Z, Luo F, Zhang L M. Research advance on molecular ecology of asymbiotic nitrogen fixation microbes. Chin J Appl Ecol, 2017, 28: 3440-3450. (in Chinese with English abstract) | |
| [29] |
Reinhold-Hurek B, Bünger W, Burbano C S, Sabale M, Hurek T. Roots shaping their microbiome: global hotspots for microbial activity. Annu Rev Phytopathol, 2015, 53: 403-424.
doi: 10.1146/annurev-phyto-082712-102342 pmid: 26243728 |
| [30] |
Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R, Ashmore M, Bustamante M, Cinderby S, Davidson E, Dentener F. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol Appl, 2010, 20: 30-59.
pmid: 20349829 |
| [31] | Liu W, Jiang L, Yang S, Wang Z, Tian R, Peng Z, Chen Y, Zhang X, Kuang J, Ling N. Critical transition of soil bacterial diversity and composition triggered by nitrogen enrichment. Ecology, 2020, 101: e03053. |
| [32] | Calderoli P A, Collavino M M, Kraemer F B, Morrás H, Aguilar O M. Analysis of nifH-RNA reveals phylotypes related to Geobacter and Cyanobacteria as important functional components of the N2-fixing community depending on depth and agriculturaluse of soil. Microbiol Open, 2017, 6: e502. |
| [33] |
Wang R, Chang Y L, Zheng W T, Zhang D, Zhang X X, Sui X H, Wang E T, Hu J Q, Zhang L Y, Chen W X. Bradyrhizobium arachidis sp. nov., isolated from effective nodules of Arachis hypogaea grown in China. Sys Appl Microbiol, 2013, 36: 101-105.
doi: 10.1016/j.syapm.2012.10.009 |
| [34] |
Indrasumunar A, Menzies N W, Dart P J. Laboratory prescreening of Bradyrhizobium japonicum for low pH, Al and Mn tolerance can be used to predict their survival in acid soils. Soil Biol Biochem, 2012, 48: 135-141.
doi: 10.1016/j.soilbio.2012.01.019 |
| [35] |
Duncan N, Menge L, Levin S, Hedin L. Facultative versus obligate nitrogen fixation strategies and their ecosystem consequences. Am Nat, 2009, 174: 465-477.
doi: 10.1086/605377 |
| [1] | 周群, 袁锐, 朱宽宇, 王志琴, 杨建昌. 不同施氮量下籼/粳杂交稻甬优2640产量和氮素吸收利用的特点[J]. 作物学报, 2022, 48(9): 2285-2299. |
| [2] | 张振博, 屈馨月, 于宁宁, 任佰朝, 刘鹏, 赵斌, 张吉旺. 施氮量对夏玉米籽粒灌浆特性和内源激素作用的影响[J]. 作物学报, 2022, 48(9): 2366-2376. |
| [3] | 张胜忠, 胡晓辉, 慈敦伟, 杨伟强, 王菲菲, 邱俊兰, 张天雨, 钟文, 于豪諒, 孙冬平, 邵战功, 苗华荣, 陈静. 基于三维模型重构的花生网纹厚度性状QTL分析[J]. 作物学报, 2022, 48(8): 1894-1904. |
| [4] | 刘昆, 黄健, 周沈琪, 张伟杨, 张耗, 顾骏飞, 刘立军, 杨建昌. 穗肥施氮量对不同穗型超级稻品种产量的影响及其机制[J]. 作物学报, 2022, 48(8): 2028-2040. |
| [5] | 白冬梅, 薛云云, 黄莉, 淮东欣, 田跃霞, 王鹏冬, 张鑫, 张蕙琪, 李娜, 姜慧芳, 廖伯寿. 不同花生品种芽期耐寒性鉴定及评价指标筛选[J]. 作物学报, 2022, 48(8): 2066-2079. |
| [6] | 徐扬, 张智猛, 丁红, 秦斐斐, 张冠初, 戴良香. 钙肥对酸性红壤花生种子萌发及种子际微生物菌群结构的调控[J]. 作物学报, 2022, 48(8): 2088-2099. |
| [7] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [8] | 李海芬, 魏浩, 温世杰, 鲁清, 刘浩, 李少雄, 洪彦彬, 陈小平, 梁炫强. 花生电压依赖性阴离子通道基因(AhVDAC)的克隆及在果针向地性反应中表达分析[J]. 作物学报, 2022, 48(6): 1558-1565. |
| [9] | 彭西红, 陈平, 杜青, 杨雪丽, 任俊波, 郑本川, 罗凯, 谢琛, 雷鹿, 雍太文, 杨文钰. 减量施氮对带状套作大豆土壤通气环境及结瘤固氮的影响[J]. 作物学报, 2022, 48(5): 1199-1209. |
| [10] | 刘嘉欣, 兰玉, 徐倩玉, 李红叶, 周新宇, 赵璇, 甘毅, 刘宏波, 郑月萍, 詹仪花, 张刚, 郑志富. 耐三唑并嘧啶类除草剂花生种质创制与鉴定[J]. 作物学报, 2022, 48(4): 1027-1034. |
| [11] | 李鑫格, 高杨, 刘小军, 田永超, 朱艳, 曹卫星, 曹强. 播期播量及施氮量对冬小麦生长及光谱指标的影响[J]. 作物学报, 2022, 48(4): 975-987. |
| [12] | 袁嘉琦, 刘艳阳, 许轲, 李国辉, 陈天晔, 周虎毅, 郭保卫, 霍中洋, 戴其根, 张洪程. 氮密处理提高迟播栽粳稻资源利用和产量[J]. 作物学报, 2022, 48(3): 667-681. |
| [13] | 丁红, 徐扬, 张冠初, 秦斐斐, 戴良香, 张智猛. 不同生育期干旱与氮肥施用对花生氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 695-703. |
| [14] | 黄莉, 陈玉宁, 罗怀勇, 周小静, 刘念, 陈伟刚, 雷永, 廖伯寿, 姜慧芳. 花生种子大小相关性状QTL定位研究进展[J]. 作物学报, 2022, 48(2): 280-291. |
| [15] | 谢呈辉, 马海曌, 许宏伟, 徐郗阳, 阮国兵, 郭峥岩, 宁永培, 冯永忠, 杨改河, 任广鑫. 施氮量对宁夏引黄灌区麦后复种糜子生长、产量及氮素利用的影响[J]. 作物学报, 2022, 48(2): 463-477. |
|