欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (6): 1312-1324.doi: 10.3724/SP.J.1006.2022.13040
崔连花**( ), 詹为民**(), 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培*(), 杨建平*(), 杨青华*()
), 詹为民**(), 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培*(), 杨建平*(), 杨青华*()
CUI Lian-Hua**(), ZHAN Wei-Min**(), YANG Lu-Hao, WANG Shao-Ci, MA Wen-Qi, JIANG Liang-Liang, ZHANG Yan-Pei*(), YANG Jian-Ping*(), YANG Qing-Hua*()
摘要:
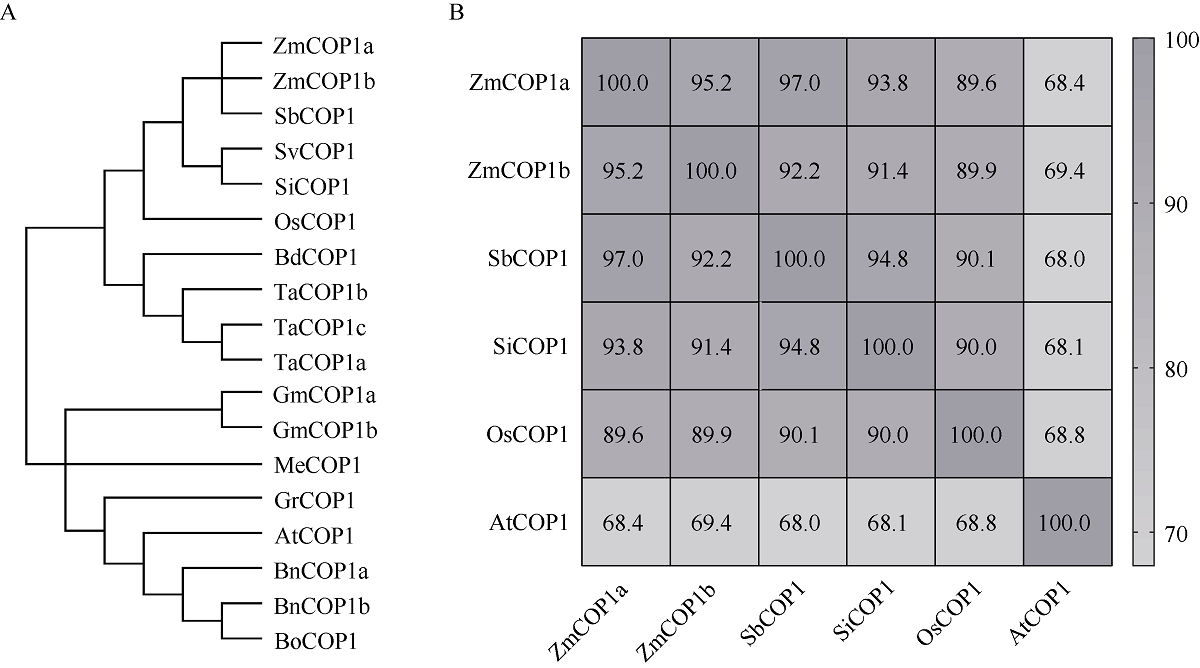
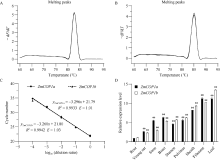
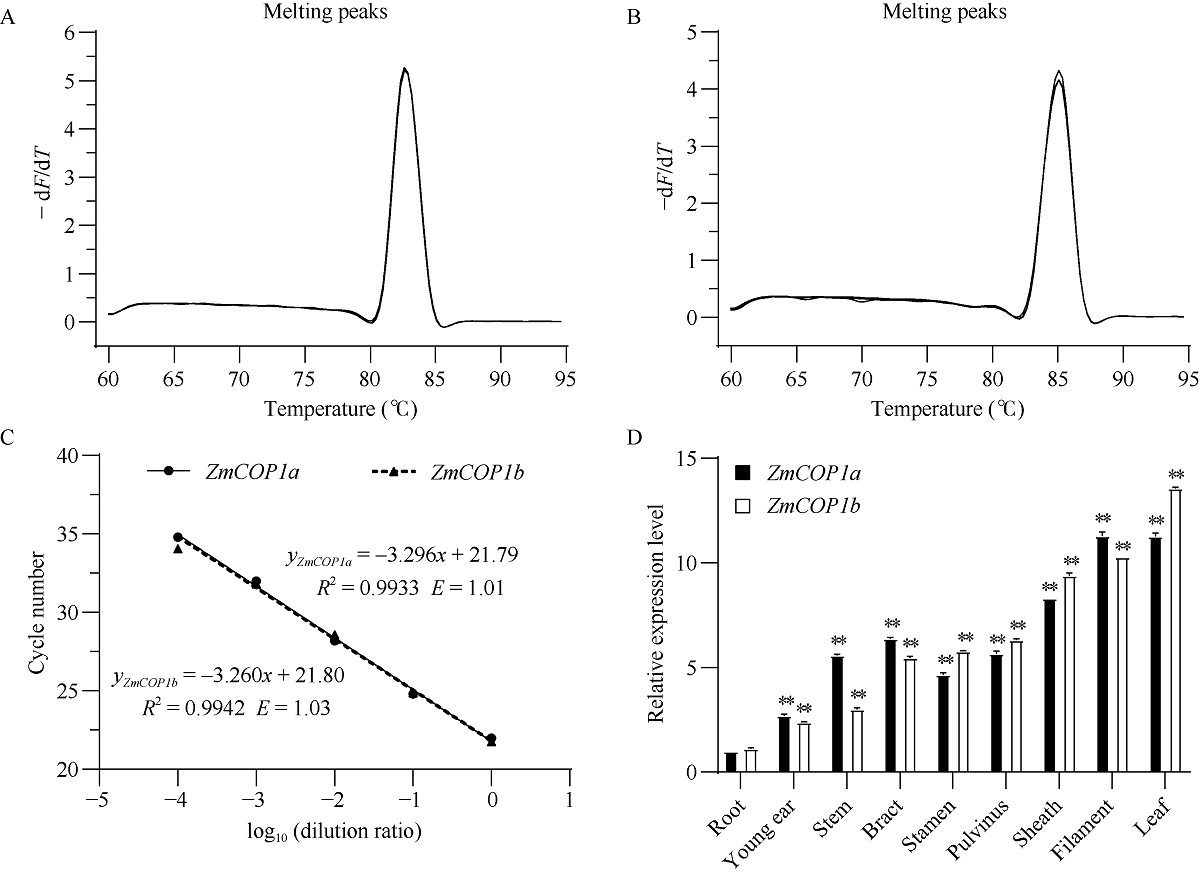
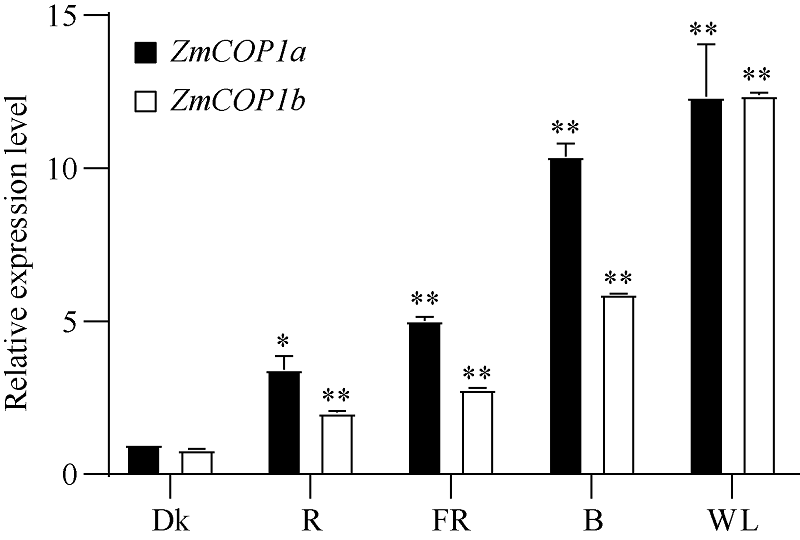
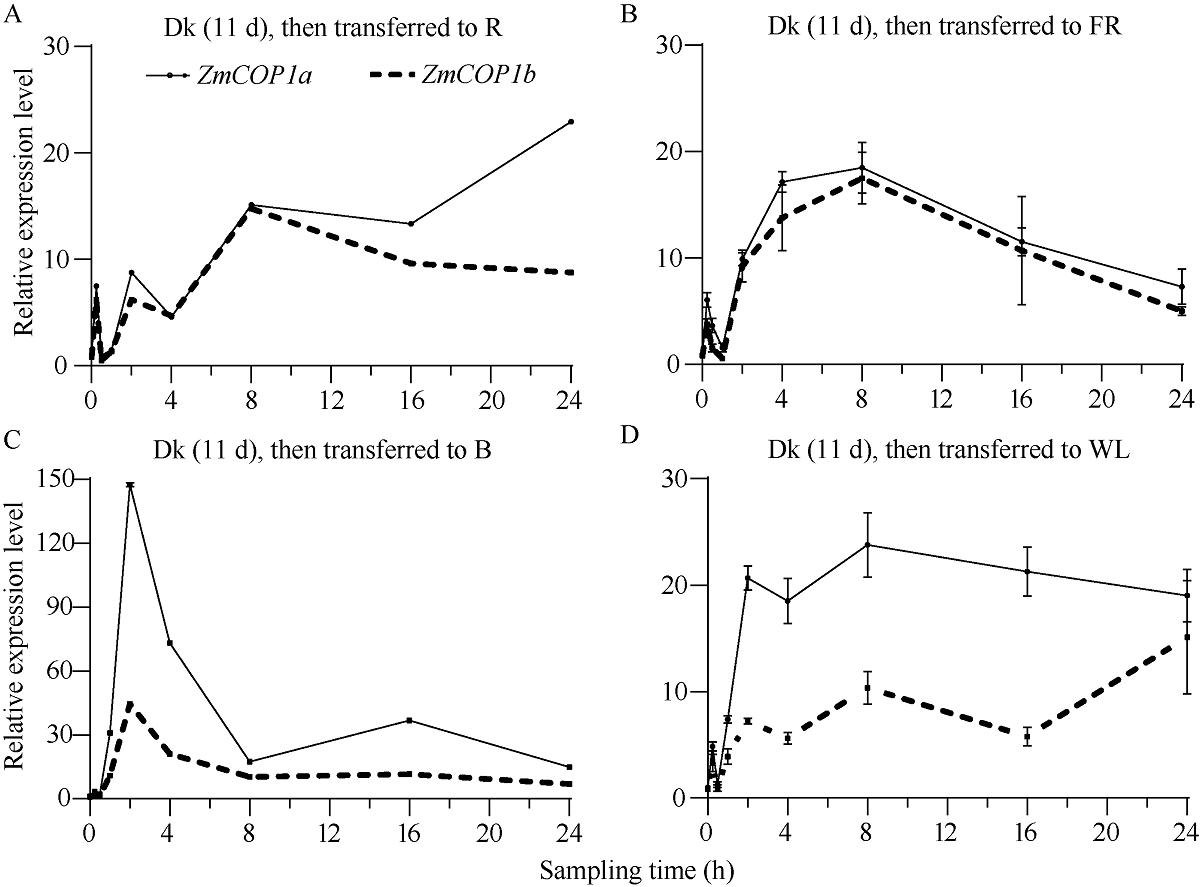
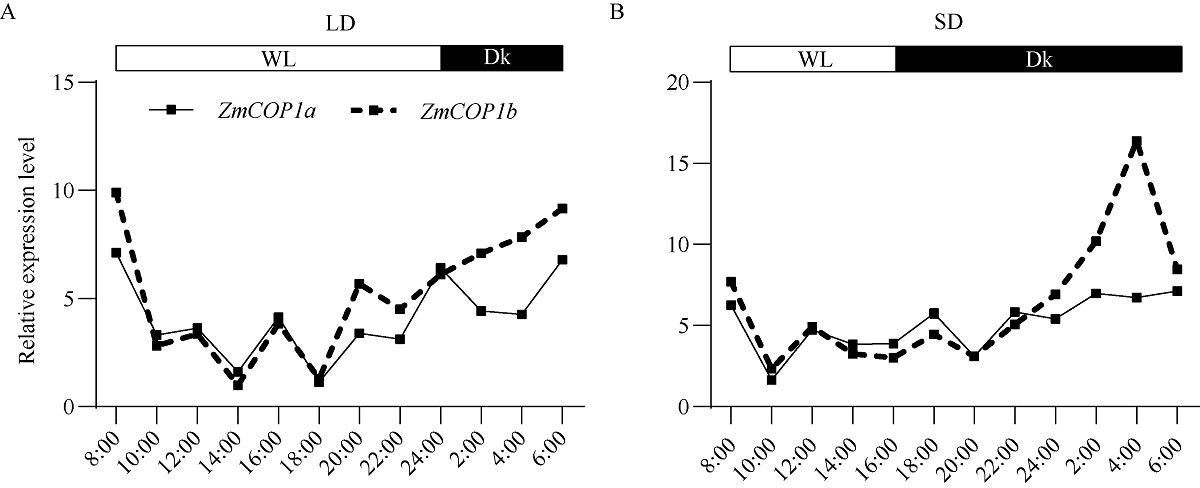
CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1)作为一种E3泛素连接酶, 能够介导拟南芥光信号转导中HY5、LAF1、HFR1和CO等正调控因子的泛素化降解, 从而调控幼苗光形态建成、花青素合成和开花等生物学过程。为了探索玉米COP1基因的功能, 本文以玉米自交系B73为研究材料, 利用反转录PCR (RT-PCR)方法克隆了2个ZmCOP1基因, 分别命名为ZmCOP1a和ZmCOP1b。通过生物信息学相关软件和网站对ZmCOP1进行理化性质分析、结构域预测和系统发育树分析, 采用实时荧光定量PCR (qRT-PCR)技术进一步分析2个ZmCOP1基因在不同组织中的转录丰度, 以及其在幼苗期对不同光质和光周期处理的响应。研究发现, ZmCOP1a和ZmCOP1b的开放阅读框(ORF)分别包含2082个和2061个核苷酸, 编码693个和686个氨基酸。玉米、水稻、高粱、谷子和拟南芥的COP1蛋白具有相同的结构域和较高的氨基酸序列一致性, 暗示它们可能具有相似的功能。2个ZmCOP1基因主要在玉米地上组织中表达, 均能迅速响应不同光质处理, 且ZmCOP1a的转录丰度普遍高于ZmCOP1b, 表明ZmCOP1a可能在不同光质处理下具有更重要的作用。在长日照和短日照条件下, ZmCOP1a和ZmCOP1b在黑暗阶段的转录丰度整体高于光照阶段, 有趣的是, 二者在光照阶段的表达模式相似, 但是在黑暗阶段, ZmCOP1b的转录丰度高于ZmCOP1a。这说明在不同光周期条件下, ZmCOP1b可能比ZmCOP1a更重要。综上这些研究结果表明, ZmCOP1a和ZmCOP1b具有功能冗余和分化。二者可以参与不同光信号通路, 可能在玉米光形态建成和花期调控中发挥重要作用。本研究为进一步探明ZmCOP1a和ZmCOP1b基因的功能及其在玉米育种中的应用提供了研究基础。
| [1] | 李合生. 现代植物生理学(第3版). 北京: 高等教育出版社, 2012. pp 287-288. |
| Li H S. Modern Plant Physiology, 3rd edn. Beijing: Higher Education Press, 2012. pp 287-288(in Chinese). | |
| [2] |
Sullivan J A, Deng X W. From seed to seed: the role of photoreceptors in Arabidopsis development. Dev Biol, 2003, 260: 289-297.
pmid: 12921732 |
| [3] |
Wang Q, Lin C T. Mechanisms of cryptochrome-mediated photoresponses in plants. Annu Rev Plant Biol, 2020, 71: 103-129.
doi: 10.1146/annurev-arplant-050718-100300 pmid: 32169020 |
| [4] |
Legris M, Ince Y Ç, Fankhauser C. Molecular mechanisms underlying phytochrome-controlled morphogenesis in plants. Nat Commun, 2019, 10: 5219.
doi: 10.1038/s41467-019-13045-0 |
| [5] |
Qian C Z, Chen Z R, Liu Q, Mao W W, Chen Y L, Tian W, Liu Y, Han J P, Ou-yang X H, Huang X. Coordinated transcriptional regulation by the UV-B photoreceptor and multiple transcription factors for plant UV-B responses. Mol Plant, 2020, 13: 777-792.
doi: 10.1016/j.molp.2020.02.015 |
| [6] |
Christie J M. Phototropin blue-light receptors. Annu Rev Plant Biol, 2007, 58: 21-45.
pmid: 17067285 |
| [7] |
Podolec R, Demarsy E, Ulm R. Perception and signaling of Ultraviolet-B radiation in plants. Annu Rev Plant Biol, 2021, 72: 793-822.
doi: 10.1146/annurev-arplant-050718-095946 pmid: 33636992 |
| [8] |
Ma L G, Gao Y, Qu L J, Chen Z L, Li J M, Zhao H Y, Deng X W. Genomic evidence for COP1 as a repressor of light-regulated gene expression and development in Arabidopsis. Plant Cell, 2002, 14: 2383-2398.
doi: 10.1105/tpc.004416 |
| [9] |
Podolec R, Ulm R. Photoreceptor-mediated regulation of the COP1/SPA E3 ubiquitin ligase. Curr Opin Plant Biol, 2018, 45: 18-25.
doi: S1369-5266(17)30208-X pmid: 29775763 |
| [10] |
Sharma A, Sharma B, Hayes S, Kerner K, Hoecker U, Jenkins G I, Franklin K A. UVR8 disrupts stabilisation of PIF5 by COP1 to inhibit plant stem elongation in sunlight. Nat Commun, 2019, 10: 4417.
doi: 10.1038/s41467-019-12369-1 |
| [11] |
Hiltbrunner A. Shedding light on the evolution of light signalling. New Phytol, 2019, 224: 1412-1414.
doi: 10.1111/nph.16175 pmid: 31587283 |
| [12] |
Han X, Huang X, Deng X W. The photomorphogenic central repressor COP1: conservation and functional diversification during evolution. Plant Commun, 2020, 1: 100044.
doi: 10.1016/j.xplc.2020.100044 |
| [13] |
Deng X W, Caspar T, Quail P H. cop1: a regulatory locus involved in light-controlled development and gene expression in Arabidopsis. Genes Dev, 1991, 5: 1172-1182.
doi: 10.1101/gad.5.7.1172 |
| [14] |
Deng X W, Matsui M, Wei N, Wagner D, Chu A M, Feldmann K A, Quail P H. COP1, an Arabidopsis regulatory gene, encodes a protein with both a zinc-binding motif and a Gβ homologous domain. Cell, 1992, 71: 791-801.
pmid: 1423630 |
| [15] |
Seo H S, Yang J Y, Ishikawa M, Bolle C, Ballesteros M L, Chua N H. LAF1 ubiquitination by COP1 controls photomorphogenesis and is stimulated by SPA1. Nature, 2003, 423: 995-999.
doi: 10.1038/nature01696 |
| [16] |
Yang J P, Lin R C, Sullivan J, Hoecker U, Liu B L, Xu L, Deng X W, Wang H Y. Light regulates COP1-mediated degradation of HFR1, a transcription factor essential for light signaling in Arabidopsis. Plant Cell, 2005, 17: 804-821.
doi: 10.1105/tpc.104.030205 |
| [17] |
Osterlund M T, Wei N, Deng X W. The roles of photoreceptor systems and the COP1-targeted destabilization of HY5 in light control of Arabidopsis seedling development. Plant Physiol, 2000, 124: 1520-1524.
pmid: 11115869 |
| [18] |
Holm M, Ma L G, Qu L J, Deng X W. Two interacting bZIP proteins are direct targets of COP1-mediated control of light- dependent gene expression in Arabidopsis. Genes Dev, 2002, 16: 1247-1259.
doi: 10.1101/gad.969702 |
| [19] |
Ling J J, Li J, Zhu D M, Deng X W. Noncanonical role of Arabidopsis COP1/SPA complex in repressing BIN2-mediated PIF3 phosphorylation and degradation in darkness. Proc Natl Acad Sci USA, 2017, 114: 3539-3544.
doi: 10.1073/pnas.1700850114 |
| [20] |
Cañibano E, Bourbousse C, García-León M, Gómez B G, Wolff L, García-Baudino C, Lozano-Durán R, Barneche F, Rubio V, Fonseca S. DET1-mediated COP1 regulation avoids HY5 activity over second-site gene targets to tune plant photomorphogenesis. Mol Plant, 2021, 14: 963-982.
doi: 10.1016/j.molp.2021.03.009 pmid: 33711490 |
| [21] |
Ponnu J. Molecular mechanisms suppressing COP1/SPA E3 ubiquitin ligase activity in blue light. Physiol Plant, 2020, 169: 418-429.
doi: 10.1111/ppl.v169.3 |
| [22] |
Yadav A, Singh D, Lingwan M, Yadukrishnan P, Masakapalli S K, Datta S. Light signaling and UV-B-mediated plant growth regulation. J Integr Plant Biol, 2020, 62: 1270-1292.
doi: 10.1111/jipb.v62.9 |
| [23] |
Oh J, Park E, Song K, Bae G, Choi G. PHYTOCHROME INTERACTING FACTOR8 inhibits phytochrome A-mediated far-red light responses in Arabidopsis. Plant Cell, 2020, 32: 186-205.
doi: 10.1105/tpc.19.00515 |
| [24] |
Ren H, Han J P, Yang P Y, Mao W W, Liu X, Qiu L L, Qian C Z, Liu Y, Chen Z R, Ou-yang X H, Chen X, Deng X W, Huang X. Two E3 ligases antagonistically regulate the UV-B response in Arabidopsis. Proc Natl Acad Sci USA, 2019, 116: 4722-4731.
doi: 10.1073/pnas.1816268116 |
| [25] |
Huang X, Ouyang X H, Yang P Y, Lau O S, Li G, Li J G, Chen H D, Deng X W. Arabidopsis FHY3 and HY5 positively mediate induction of COP1 transcription in response to photomorphogenic UV-B light. Plant Cell, 2012, 24: 4590-4606.
doi: 10.1105/tpc.112.103994 |
| [26] |
Tavridou E, Pireyre M, Ulm R. Degradation of the transcription factors PIF4 and PIF5 under UV-B promotes UVR8-mediated inhibition of hypocotyl growth in Arabidopsis. Plant J, 2020, 101: 507-517.
doi: 10.1111/tpj.14556 |
| [27] |
Yu Y W, Wang J, Shi H, Gu J T, Dong J G, Deng X W, Huang R F. Salt stress and ethylene antagonistically regulate nucleocytoplasmic partitioning of COP1 to control seed germination. Plant Physiol, 2016, 170: 2340-2350.
doi: 10.1104/pp.15.01724 |
| [28] |
Shi H, Liu R L, Xue C, Shen X, Wei N, Deng X W, Zhong S W. Seedlings transduce the depth and mechanical pressure of covering soil using COP1 and ethylene to regulate EBF1/EBF2 for soil emergence. Curr Biol, 2016, 26: 139-149.
doi: 10.1016/j.cub.2015.11.053 |
| [29] |
Tanaka N, Itoh H, Sentoku N, Kojima M, Sakakibara H, Izawa T, Itoh J I, Nagato Y. The COP1 ortholog PPS regulates the juvenile-adult and vegetative-reproductive phase changes in rice. Plant Cell, 2011, 23: 2143-2154.
doi: 10.1105/tpc.111.083436 |
| [30] |
Liu L J, Zhang Y C, Li Q H, Sang Y, Mao J, Lian H L, Wang L, Yang H Q. COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis. Plant Cell, 2008, 20: 292-306.
doi: 10.1105/tpc.107.057281 pmid: 18296627 |
| [31] |
Zuo Z C, Liu H T, Liu B, Liu X M, Lin C T. Blue light-dependent interaction of CRY2 with SPA1 regulates COP1 activity and floral initiation in Arabidopsis. Curr Biol, 2011, 21: 841-847.
doi: 10.1016/j.cub.2011.03.048 |
| [32] |
Kang C Y, Lian H L, Wang F F, Huang J R, Yang H Q. Cryptochromes, phytochromes, and COP1 regulate light-controlled stomatal development in Arabidopsis. Plant Cell, 2009, 21: 2624-2641.
doi: 10.1105/tpc.109.069765 |
| [33] |
Zhao C Z, Wang P C, Si T, Hsu C C, Wang L, Zayed O, Yu Z P, Zhu Y F, Dong J, Tao W A, Zhu J K. MAP Kinase cascades regulate the cold response by modulating ICE1 protein stability. Dev Cell, 2017, 43: 618-629.
doi: 10.1016/j.devcel.2017.09.024 |
| [34] |
Wei H B, Kong D X, Yang J, Wang H Y. Light regulation of stomatal development and patterning: shifting the paradigm from Arabidopsis to grasses. Plant Commun, 2020, 1: 100030.
doi: 10.1016/j.xplc.2020.100030 |
| [35] |
Lee J H, Jung J H, Park C M. Light inhibits COP1-mediated degradation of ICE transcription factors to induce stomatal development in Arabidopsis. Plant Cell, 2017, 29: 2817-2830.
doi: 10.1105/tpc.17.00371 |
| [36] |
Chen Q B, Bai L, Wang W J, Shi H Z, Botella J R, Zhan Q D, Liu K, Yang H Q, Song C P. COP1 promotes ABA-induced stomatal closure by modulating the abundance of ABI/HAB and AHG3 phosphatases. New Phytol, 2021, 229: 2035-2049.
doi: 10.1111/nph.v229.4 |
| [37] | Lu Y F, Hao S X, Liu N, Bu Y F, Yang S L, Yao Y C. Light affects anthocyanin biosynthesis via transcriptional regulation of COP1 in the ever-red leaves of crabapple M. cv. ‘Royalty’. Brazilian J Bot, 2016, 39: 659-667. |
| [38] |
Wu M, Si M, Li X Y, Song L Y, Liu J L, Zhai R, Cong L, Yue R R, Yang C Q, Ma F W, Xu L F, Wang Z G. PbCOP1.1 contributes to the negative regulation of anthocyanin biosynthesis in pear. Plants (Basel), 2019, 8: 39.
doi: 10.3390/plants8020039 |
| [39] | 蒋明敏. 光信号诱导茄子花青素合成的分子机制研究. 上海交通大学博士学位论文, 上海, 2016. |
| Jiang M M. The Molecular Mechanism of Light Signal Induced Anthocyanin Biosynthesis in Solanum melongena. PhD Dissertation of Shanghai Jiao Tong University, Shanghai, China, 2016 (in Chinese with English abstract). | |
| [40] |
Hao X L, Zhong Y J, Nützmann H W, Fu X Q, Yan T X, Shen Q, Chen M H, Ma Y N, Zhao J Y, Osbourn A, Li L, Tang K X. Light-induced artemisinin biosynthesis is regulated by the bZIP transcription factor AaHY5 in Artemisia annua. Plant Cell Physiol, 2019, 60: 1747-1760.
doi: 10.1093/pcp/pcz084 |
| [41] |
Rajeevan M S, Ranamukhaarachchi D G, Vernon S D, Unger E R. Use of real-time quantitative PCR to validate the results of cDNA array and differential display PCR technologies. Methods, 2001, 25: 443-451.
pmid: 11846613 |
| [42] |
蒋可人, 马峥, 郑航, 刘小军. 转录组与蛋白质组整合分析在生物学研究中的应用. 生物技术通报, 2018, 34(12):50-55.
doi: 10.13560/j.cnki.biotech.bull.1985.2017-0929 |
| Jiang K R, Ma Z, Zheng H, Liu X J. Review on the application of integrated transcriptome and proteome analysis in biology. Biotechnol Bull, 2018, 34(12):50-55 (in Chinese with English abstract). | |
| [43] |
Liu Y S, Beyer A, Aebersold R. On the dependency of cellular protein levels on mRNA abundance. Cell, 2016, 165: 535-550.
doi: 10.1016/j.cell.2016.03.014 |
| [44] |
Torii K U, McNellis T W, Deng X W. Functional dissection of Arabidopsis COP1 reveals specific roles of its three structural modules in light control of seedling development. EMBO J, 1998, 17: 5577-5587.
pmid: 9755158 |
| [45] |
Zhu D M, Maier A, Lee J H, Laubinger S, Saijo Y, Wang H Y, Qu L J, Hoecker U, Deng X W. Biochemical characterization of Arabidopsis complexes containing CONSTITUTIVELY PHOTOMORPHOGENIC1 and SUPPRESSOR OF PHYA proteins in light control of plant development. Plant Cell, 2008, 20: 2307-2323.
doi: 10.1105/tpc.107.056580 |
| [46] |
Holm M, Hardtke C S, Gaudet R, Deng X W. Identification of a structural motif that confers specific interaction with the WD40 repeat domain of Arabidopsis COP1. EMBO J, 2001, 20: 118-127.
pmid: 11226162 |
| [47] |
Jang I C, Yang J Y, Seo H S, Chua N H. HFR1 is targeted by COP1 E3 ligase for post-translational proteolysis during phytochrome A signaling. Genes Dev, 2005, 19: 593-602.
doi: 10.1101/gad.1247205 |
| [48] | Huai J L, Jing Y J, Lin R C. Functional analysis of ZmCOP1 and ZmHY5 reveals conserved light signaling mechanism in maize and Arabidopsis. Physiol Plant, 2020, 169: 369-379. |
| [49] |
Liu B, Zuo Z C, Liu H T, Liu X M, Lin C T. Arabidopsis cryptochrome 1 interacts with SPA1 to suppress COP1 activity in response to blue light. Genes Dev, 2011, 25: 1029-1034.
doi: 10.1101/gad.2025011 |
| [50] |
Liu H T, Liu B, Zhao C X, Pepper M, Lin C T. The action mechanisms of plant cryptochromes. Trends Plant Sci, 2011, 16: 684-691.
doi: 10.1016/j.tplants.2011.09.002 |
| [51] | 李仕铭, 周增, 涂敏, 叶肖肖, 林辰涛, 左泽乘. 拟南芥隐花色素CRY光信号通路的研究进展. 分子植物育种, 2018, 16: 4444-4452. |
| Li S M, Zhou Z, Tu M, Ye X X, Lin C T, Zuo Z C. Research progress of photosignal pathway of cryptochrome in Arabidopsis thaliana. Mol Plant Breed, 2018, 16: 4444-4452 (in Chinese with English abstract). | |
| [52] | 周婷婷. 甜高粱隐花色素CRYPTOCHROME 1a和CRYPTOCHROME 1b的功能、信号转导机制及胁迫响应分析. 吉林大学博士学位论文, 吉林长春, 2017. |
| Zhou T T. Function, Signaling Mechanism and Stress Response of the CRYPTOCHROME 1a and CRYPTOCHROME 1b in Sweet Sorghum. PhD Dissertation of Jilin University, Changchun, Jilin, China, 2017 (in Chinese with English abstract). | |
| [53] | Vogel C, Marcotte E M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat Rev Genet, 2012, 13: 227-232. |
| [54] |
Wang B B, Lin Z C, Li X, Zhao Y P, Zhao B B, Wu G X, Ma X J, Wang H, Xie Y R, Li Q Q, Song G S, Kong D X, Zheng Z G, Wei H B, Shen R X, Wu H, Chen C X, Meng Z D, Wang T Y, Li Y, Li X H, Chen Y H, Lai J S, Hufford M B, Ross-Ibarra J, He H, Wang H Y. Genome-wide selection and genetic improvement during modern maize breeding. Nat Genet, 2020, 52: 565-571.
doi: 10.1038/s41588-020-0616-3 |
| [55] |
Ducrocq S, Giauffret C, Madur D, Combes V, Dumas F, Jouanne S, Coubriche D, Jamin P, Moreau L, Charcosset A. Fine mapping and haplotype structure analysis of a major flowering time quantitative trait locus on maize chromosome 10. Genetics, 2009, 183: 1555-1563.
doi: 10.1534/genetics.109.106922 |
| [56] |
Zhou Z Q, Zhang C S, Zhou Y, Hao Z F, Wang Z H, Zeng X, Di H, Li M S, Zhang D G, Yong H J, Zhang S H, Weng J F, Li X H. Genetic dissection of maize plant architecture with an ultra-high density bin map based on recombinant inbred lines. BMC Genom, 2016, 17: 178.
doi: 10.1186/s12864-016-2555-z |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [3] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [4] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [5] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [6] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [7] | 周慧文, 丘立杭, 黄杏, 李强, 陈荣发, 范业赓, 罗含敏, 闫海锋, 翁梦苓, 周忠凤, 吴建明. 甘蔗赤霉素氧化酶基因ScGA20ox1的克隆及功能分析[J]. 作物学报, 2022, 48(4): 1017-1026. |
| [8] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [9] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [10] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [11] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [12] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [13] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [14] | 张倩, 韩本高, 张博, 盛开, 李岚涛, 王宜伦. 控失尿素减施及不同配比对夏玉米产量及氮肥效率的影响[J]. 作物学报, 2022, 48(1): 180-192. |
| [15] | 苏达, 颜晓军, 蔡远扬, 梁恬, 吴良泉, MUHAMMAD AtifMuneer, 叶德练. 磷肥对甜玉米籽粒植酸和锌有效性的影响[J]. 作物学报, 2022, 48(1): 203-214. |
|
||