欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (4): 966-977.doi: 10.3724/SP.J.1006.2023.21023
周宾寒1( ), 杨竹1, 王书平1, 方正武1, 胡赞民2, 徐兆师3, 张迎新1,*()
), 杨竹1, 王书平1, 方正武1, 胡赞民2, 徐兆师3, 张迎新1,*()
ZHOU Bin-Han1(), YANG Zhu1, WANG Shu-Ping1, FANG Zheng-Wu1, HU Zan-Min2, XU Zhao-Shi3, ZHANG Ying-Xin1,*()
摘要:
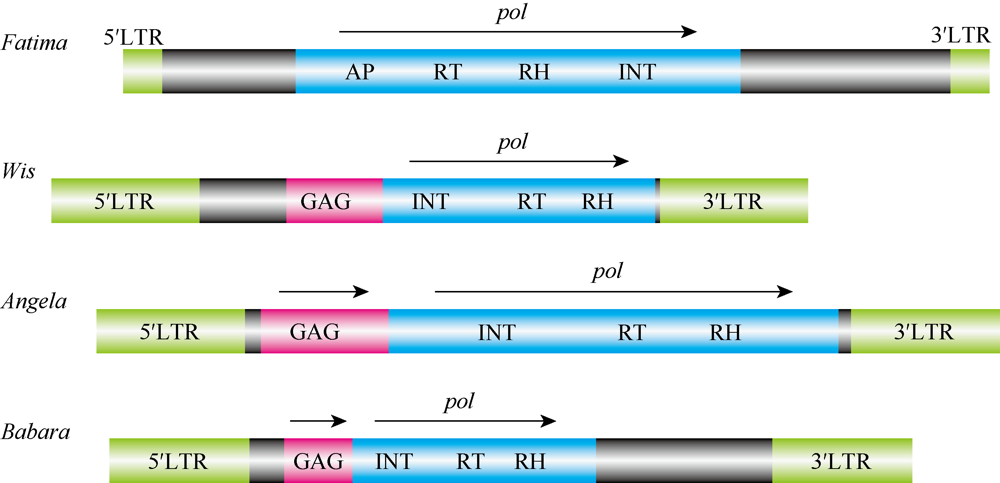
LTR (长末端重复, long terminal repeat)反转录转座子占小麦基因组的60%以上, 筛选小麦基因组中具有转座活性的LTR反转录转座子, 并分析其在非生物胁迫下的响应, 对研究反转录转座子在小麦抗逆境胁迫中的作用具有重要意义。本研究通过生物信息学分析, 从转座子数据库(TREP database)中筛选出4个具有完整结构的LTR反转录转座子Fatima、Wis、Angela和Babara; 同时利用实时荧光定量PCR (qRT-PCR)、甲基化特异PCR (methylmion specific PCR, MSP)和转座子展示(transposon display, TD)技术分别分析了它们在盐、ABA、H2O2和干旱等处理的小麦幼苗期(二叶一心)叶和根中的表达水平、甲基化水平和转座活性变化。结果表明, 这4个反转录转座子在正常条件下均存在基础水平的转录, 并且能够响应上述4种胁迫而发生转录水平的变化, 且在相同胁迫条件下表达水平变化趋势一致。Fatima、Angela和Babara在非生物胁迫处理下表达水平的提高与其甲基化水平的降低有关, Wis则相反。反转录转座子LTR序列含有胁迫响应顺式作用元件, 但在非生物胁迫条件下顺式作用元件对这4个反转录转座子的调控作用不显著。与叶相比, 这4个反转录转座子在根中对胁迫的响应程度更高, 且在盐和ABA处理下转座活性更强。本研究将有助于进一步揭示LTR反转录转座子对非生物胁迫的响应规律, 为进一步研究利用反转录转座子进行小麦抗逆育种的遗传改良积累资料。
| [1] |
Feschotte C, Jiang N, Wessler S R. Plant transposable elements: where genetics meets genomics. Nat Rev Genet, 2002, 3: 329-341.
doi: 10.1038/nrg793 pmid: 11988759 |
| [2] |
Hou J, Lu D, Mason A S, Li B, Xiao M, An S, Fu D. Non-coding RNAs and transposable elements in plant genomes: emergence, regulatory mechanisms and roles in plant development and stress responses. Planta, 2019, 250: 23-40.
doi: 10.1007/s00425-019-03166-7 pmid: 30993403 |
| [3] | Ning J. Plant transposable Elements:Beyond Insertions and Interruptions. New York: Springer-Verlag, 2018. |
| [4] |
Roquis D, Robertson M, Yu L, Thieme M, Julkowska M, Bucher E. Genomic impact of stress-induced transposable element mobility in Arabidopsis. Nucleic Acids Res, 2021, 49: 10431-10447.
doi: 10.1093/nar/gkab828 |
| [5] |
Wicker T, Gundlach H, Spannagl M, Uauy C, Borrill P, Ramírez-González R H, De Oliveira R. International Wheat Genome Sequencing Consortium, Mayer Kfx, Paux E, Choulet F. Impact of transposable elements on genome structure and evolution in bread wheat. Genome Biol, 2018, 19: 103.
doi: 10.1186/s13059-018-1479-0 pmid: 30115100 |
| [6] |
Alonge M, Shumate A, Puiu D, Zimin A V, Salzberg S L. Chromosome-scale assembly of the bread wheat genome reveals thousands of additional gene copies. Genetics, 2020, 216: 599-608.
doi: 10.1534/genetics.120.303501 pmid: 32796007 |
| [7] |
Hirsch C D, Springer N M. Transposable element influences on gene expression in plants. Biochim Biophys Acta Gene Regul Mech, 2017, 1860: 157-165.
doi: 10.1016/j.bbagrm.2016.05.010 |
| [8] |
Piégu B, Bire S, Arensburger P, Bigot Y. A survey of transposable element classification systems: a call for a fundamental update to meet the challenge of their diversity and complexity. Mol Phylogenet Evol, 2015, 86: 90-109.
doi: 10.1016/j.ympev.2015.03.009 |
| [9] |
Ramakrishnan M, Satish L, Kalendar R, Narayanan M, Kandasamy S, Sharma A, Emamverdian A, Wei Q, Zhou M. The dynamism of transposon methylation for plant development and stress adaptation. Int J Mol Sci, 2021, 22: 11387.
doi: 10.3390/ijms222111387 |
| [10] |
Galindo-González L, Sarmiento F, Quimbaya M A. Shaping plant adaptability, genome structure and gene expression through transposable element epigenetic control: focus on methylation. Agronomy, 2018, 8: 180.
doi: 10.3390/agronomy8090180 |
| [11] |
Saze H. Epigenetic regulation of intragenic transposable elements: a two-edged sword. J Biochem, 2018, 164: 323-328.
doi: 10.1093/jb/mvy060 pmid: 30010918 |
| [12] |
Yates-Stewart A D, Daron J, Wijeratne S, Shahid S, Edgington H A, Slotkin R K, Michel A. Soybean aphids adapted to host-plant resistance by down regulating putative effectors and up regulating transposable elements. Insect Biochem Mol Biol, 2020, 121: 103363.
doi: 10.1016/j.ibmb.2020.103363 |
| [13] |
Kimura Y, Tosa Y, Shimada S, Sogo R, Kusaba M, Sunaga T, Betsuyaku S, Eto Y, Nakayashiki H, Mayama S. OARE-1, a Ty1-copia retrotransposon in oat activated by abiotic and biotic stresses. Plant Cell Physiol, 2001, 42: 1345-1354.
doi: 10.1093/pcp/pce171 pmid: 11773527 |
| [14] |
Woodrow P, Pontecorvo G, Fantaccione S, Fuggi A, Kafantaris I, Parisi D, Carillo P. Polymorphism of a new Ty1-copia retrotransposon in durum wheat under salt and light stresses. Theor Appl Genet, 2010, 121: 311-322.
doi: 10.1007/s00122-010-1311-z pmid: 20237753 |
| [15] |
Roquis D, Robertson M, Yu L, Thieme M, Julkowska M, Bucher E. Genomic impact of stress-induced transposable element mobility in Arabidopsis. Nucleic Acids Res, 2021, 49: 10431-10447.
doi: 10.1093/nar/gkab828 |
| [16] |
Zhao Y, Wu L, Fu Q, Wang D, Li J, Yao B, Yu S, Jiang L, Qian J, Zhou X, Han L, Zhao S, Ma C, Zhang Y, Luo C, Dong Q, Li S, Zhang L, Jiang X, Li Y, Luo H, Li K, Yang J, Luo Q, Li L, Peng S, Huang H, Zuo Z, Liu C, Wang L, Li C, He X, Friml J, Du Y. INDITTO2 transposon conveys auxin-mediated DRO1 transcription for rice drought avoidance. Plant Cell Environ, 2021, 44: 1846-1857.
doi: 10.1111/pce.14029 |
| [17] |
Marcussen T, Sandve S R, Heier L, Spannagl M, Pfeifer M, International Wheat Genome Sequencing Consortium, Jakobsen K S, Wulff B B, Steuernagel B, Mayer K F, Olsen O A. Ancient hybridizations among the ancestral genomes of bread wheat. Science, 2014, 345: 1250092.
doi: 10.1126/science.1250092 |
| [18] | 郭靖宇. 小麦中国春全基因组转座元件的特征分析. 河南大学硕士学位论文, 河南郑州, 2020. |
| Guo J Y. Genome-wide Analysis of Transposable Elements in Bread Wheat. MS Thesis of Henan University, Zhengzhou, Henan, China, 2020. (in Chinese with English abstract) | |
| [19] |
Nian L L, Liu X L, Yang Y B, Zhu X L, Yi X F, Haider F U. Genome-wide identification, phylogenetic, and expression analysis under abiotic stress conditions of LIM gene family in Medicago sativa L. PLoS One, 2021, 16: e0252213.
doi: 10.1371/journal.pone.0252213 |
| [20] |
Mani B, Agarwal M, Katiyar-Agarwal S. Comprehensive expression profiling of rice Tetraspanin genes reveals diverse roles during development and abiotic stress. Front Plant Sci, 2015, 6: 1088.
doi: 10.3389/fpls.2015.01088 pmid: 26697042 |
| [21] |
Zhou Y B, Li Y, Qi X L, Liu R B, Dong J H, Jing W H, Guo M M, Si Q L, Xu Z S, Li L C, Wang C S, Cheng X G, Ma Y Z, Chen M. Overexpression of V-type H+pyrophosphatase gene EdVP1 from Elymus dahuricus increases yield and potassium uptake of transgenic wheat under low potassium conditions. Sci Rep, 2020, 10: 5020.
doi: 10.1038/s41598-020-62052-5 |
| [22] |
Mosoni P, Chaucheyras-Durand F, Béra-Maillet C, Forano E. Quantification by real-time PCR of cellulolytic bacteria in the rumen of sheep after supplementation of a forage diet with readily fermentable carbohydrates: effect of a yeast additive. J Appl Microbiol, 2007, 103: 2676-2685.
pmid: 18045448 |
| [23] |
Feldman M, Levy A A. Genome evolution due to allopolyploidization in wheat. Genetics, 2012, 192: 763-774.
doi: 10.1534/genetics.112.146316 pmid: 23135324 |
| [24] |
Mcclelland M, Nelson M, Raschke E. Effect of site-specific modification on restriction endonucleases and DNA modification methyltransferases. Nucleic Acids Res, 1994, 22: 3640-3659.
pmid: 7937074 |
| [25] |
Pan Y P, Bo K L, Cheng Z H, Weng Y Q. The loss-of-function GLABROUS 3 mutation in cucumber is due to LTR- retrotransposon insertion in a class IV HD-ZIP transcription factor gene CsGL3 that is epistatic over CsGL1. BMC Plant Biol, 2015, 15: 302.
doi: 10.1186/s12870-015-0693-0 |
| [26] |
Simpson S D, Nakashima K, Narusaka Y, Seki M, Shinozaki K, Yamaguchi-Shinozaki K. Two different novel cis-acting elements of erd1, a clpA homologous Arabidopsis gene function in induction by dehydration stress and dark-induced senescence. Plant J, 2003, 33: 259-270.
doi: 10.1046/j.1365-313x.2003.01624.x pmid: 12535340 |
| [27] |
Hattori T, Terada T, Hamasuna S. Regulation of the Osem gene by abscisic acid and the transcriptional activator VP1: analysis of cis-acting promoter elements required for regulation by abscisic acid and VP1. Plant J, 1995, 7: 913-925.
pmid: 7599651 |
| [28] |
Busk P K, Jensen A B, Pagès M. Regulatory elements in vivo in the promoter of the abscisic acid responsive gene rab17 from maize. Plant J, 1997, 11: 1285-1295.
pmid: 9225468 |
| [29] |
Baker S S, Wilhelm K S, Thomashow M F. The 5'-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold-, drought- and ABA-regulated gene expression. Plant Mol Biol, 1994, 24: 701-713.
doi: 10.1007/BF00029852 pmid: 8193295 |
| [30] |
Hartmann U, Sagasser M, Mehrtens F, Stracke R, Weisshaar B. Differential combinatorial interactions of cis-acting elements recognized by R2R3-MYB, BZIP, and BHLH factors control light-responsive and tissue-specific activation of phenylpropanoid biosynthesis genes. Plant Mol Biol, 2005, 57: 155-171.
doi: 10.1007/s11103-004-6910-0 pmid: 15821875 |
| [31] |
Abe H, Urao T, Ito T, Seki M, Shinozaki K, Yamaguchi- Shinozaki K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell, 2003, 15: 63-78.
doi: 10.1105/tpc.006130 |
| [32] |
Hirochika H. Activation of tobacco retrotransposons during tissue culture. EMBO J, 1993, 12: 2521-2528.
doi: 10.1002/j.1460-2075.1993.tb05907.x pmid: 8389699 |
| [33] |
Wendel J F, Wessler S R. Retrotransposon-mediated genome evolution on a local ecological scale. Proc Natl Acad Sci USA, 2000, 97: 6250-6252.
pmid: 10841529 |
| [34] | Aliyev A T, Lobianco F, Krager K J, Aykin-Burns N. Assessment of cellular oxidation using a subcellular compartment-specific redox-sensitive green fluorescent protein. J Vis Exp, 2020, 160: 10.3791/61229. |
| [35] |
Galindo-González L, Mhiri C, Deyholos M K, Grandbastien M A. LTR-retrotransposons in plants: engines of evolution. Gene, 2017, 626: 14-25.
doi: S0378-1119(17)30322-0 pmid: 28476688 |
| [36] |
Georgiev S, Dekova T, Atanassov I, Angelova Z, Dimitrova A, Mirkova V, Stoilov L. Transposable elements in wheat and Triticale sphaerococcum mutant forms. Biotechnol Biotechnol Equip, 2000, 14: 25-32.
doi: 10.1080/13102818.2000.10819057 |
| [37] | Gu Y Q, Crossman C, Kong X, Luo M, You F M, Coleman-Derr D, Dubcovsky J, Anderson O D. Genomic organization of the complex α-gliadin gene loci in wheat. J Appl Genet, 2004, 109: 648-657. |
| [38] |
Kashkush K, Feldman M, Levy A A. Transcriptional activation of retrotransposons alters the expression of adjacent genes in wheat. Nat Genet, 2003, 33: 102-106.
doi: 10.1038/ng1063 pmid: 12483211 |
| [39] |
Murphy G, Lucas H, Moore G, Flavell R. Sequence analysis of WIS-2-1A, a retrotransposon-like element from wheat. Plant Mol Biol, 1992, 20: 991-995.
doi: 10.1007/BF00027169 pmid: 1334439 |
| [40] |
Roquis D, Robertson M, Yu L, Thieme M, Julkowska M, Bucher E. Genomic impact of stress-induced transposable element mobility in Arabidopsis. Nucleic Acids Res, 2021, 49: 10431-10447.
doi: 10.1093/nar/gkab828 |
| [41] |
Wicker T, Stein N, Albar L, Feuillet C, Schlagenhauf E, Keller B. Analysis of a contiguous 211 kb sequence in diploid wheat (Triticum monococcum L.) reveals multiple mechanisms of genome evolution. Plant J, 2001, 26: 307-316.
pmid: 11439119 |
| [42] |
Gu Y Q, Crossman C, Kong X, Luo M, You F M, Coleman-Derr D, Dubcovsky J, Anderson O D. Genomic organization of the complex alpha-gliadin gene loci in wheat. Theor Appl Genet, 2004, 109: 648-657.
pmid: 15103408 |
| [43] |
Li W, Zhang P, Fellers J P, Friebe B, Gill B S. Sequence composition, organization, and evolution of the core Triticeae genome. Plant J, 2004, 40: 500-511.
doi: 10.1111/j.1365-313X.2004.02228.x |
| [44] |
Lee S C, Martienssen R A. Regulation of retrotransposition in Arabidopsis. Biochem Soc Trans, 2021, 49: 2241-2251.
doi: 10.1042/BST20210337 |
| [45] | Merkulov P, Omarov M, Gvaramiya S, Kirov I. Detection of active LTR retrotransposons via eccDNA analysis in Helianthus annuus L., Arabidopsis thaliana and triticale. In: Salina E A, ed. Plant Genetics, Genomics, Bioinformatics, and Biotechnology. Novosibirsk, Russia: The 6th International Scientific Conference, 2021 (in Russian and English). |
| [1] | 刘佳, 龚方仪, 刘亚西, 颜泽洪, 钟晓英, 陈厚霖, 黄林, 伍碧华. 野生二粒小麦主要农艺特性融入普通小麦的全基因组关联分析[J]. 作物学报, 2023, 49(5): 1184-1196. |
| [2] | 贾玉库, 高宏欢, 冯健超, 郝紫瑞, 王晨阳, 谢迎新, 郭天财, 马冬云. 小麦G2-like转录因子家族基因鉴定与表达模式分析[J]. 作物学报, 2023, 49(5): 1410-1425. |
| [3] | 张晓, 陆成彬, 江伟, 张勇, 吕国锋, 吴宏亚, 王朝顺, 李曼, 吴素兰, 高德荣. 弱筋小麦育种品质选择指标及亲本组配原则[J]. 作物学报, 2023, 49(5): 1282-1291. |
| [4] | 孙现军, 姜奇彦, 胡正, 李宏博, 庞斌双, 张风廷, 张胜全, 张辉. 小麦种质资源苗期耐盐性鉴定评价[J]. 作物学报, 2023, 49(4): 1132-1139. |
| [5] | 孙全喜, 苑翠玲, 牟艺菲, 闫彩霞, 赵小波, 王娟, 王奇, 孙慧, 李春娟, 单世华. 花生SWEET基因全基因组鉴定及表达分析[J]. 作物学报, 2023, 49(4): 938-954. |
| [6] | 张金鑫, 葛均筑, 马玮, 丁在松, 王新兵, 李从锋, 周宝元, 赵明. 华北平原冬小麦-夏玉米种植体系周年水分高效利用研究进展[J]. 作物学报, 2023, 49(4): 879-892. |
| [7] | 朱治, 李龙, 李超男, 毛新国, 郝晨阳, 朱婷, 王景一, 常建忠, 景蕊莲. 小麦转录因子TaMYB5-3B与株高和千粒重相关[J]. 作物学报, 2023, 49(4): 906-916. |
| [8] | 徐子寅, 于晓玲, 邹良平, 赵平娟, 李文彬, 耿梦婷, 阮孟斌. 木薯MYB转录因子基因MeMYB60表达特征分析及其互作蛋白筛选[J]. 作物学报, 2023, 49(4): 955-965. |
| [9] | 杨斌, 乔玲, 赵佳佳, 武棒棒, 温宏伟, 张树伟, 郑兴卫, 郑军. 小麦旗叶叶绿素含量的QTL定位及验证[J]. 作物学报, 2023, 49(3): 744-754. |
| [10] | 王雪, 谷淑波, 林祥, 王威雁, 张保军, 朱俊科, 王东. 微喷补灌水肥一体化对冬小麦产量及水分和氮素利用效率的影响[J]. 作物学报, 2023, 49(3): 784-794. |
| [11] | 高春华, 冯波, 李国芳, 李宗新, 李升东, 曹芳, 慈文亮, 赵海军. 施氮量对花后高温胁迫下冬小麦籽粒淀粉合成的影响[J]. 作物学报, 2023, 49(3): 821-832. |
| [12] | 邓照, 蒋环琪, 程丽沙, 刘睿, 黄敏, 李曼菲, 杜何为. 利用WGCNA鉴定玉米非生物胁迫相关基因共表达网络[J]. 作物学报, 2023, 49(3): 672-686. |
| [13] | 黄震, 吴启境, 陈灿妮, 吴霞, 曹珊, 张辉, 岳娇, 胡亚丽, 罗登杰, 李赟, 廖长君, 李茹, 陈鹏. 钙调素基因(HcCaM7)及其蛋白乙酰化修饰参与红麻响应非生物胁迫的作用[J]. 作物学报, 2023, 49(2): 402-413. |
| [14] | 刘方方, 万映秀, 曹文昕, 李耀, 张琪琪, 李炎, 张平治. 小麦倒春寒抗性评价方法研究[J]. 作物学报, 2023, 49(2): 438-446. |
| [15] | 孟雨, 田文仲, 温鹏飞, 丁志强, 张学品, 贺利, 段剑钊, 刘万代, 郭天财, 冯伟. 基于不同发育阶段协同的小麦品种抗旱性综合评判[J]. 作物学报, 2023, 49(2): 570-582. |
|