欢迎访问作物学报,今天是
作物学报 ›› 2018, Vol. 44 ›› Issue (03): 397-404.doi: 10.3724/SP.J.1006.2018.00397
梁云飞1,*, 张林成1,*, 蒲全明2, 雷镇泽1, 施松梅1, 姜宇鹏1, 任雪松1, 高启国1,*( )
)
Yun-Fei LIANG1,**, Lin-Cheng ZHANG1,**, Quan-Ming PU2, Zhen-Ze LEI1, Song-Mei SHI1, Yu-Peng JIANG1, Xue-Song REN1, Qi-Guo GAO1,*()
摘要:
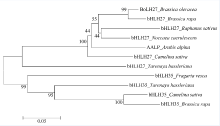
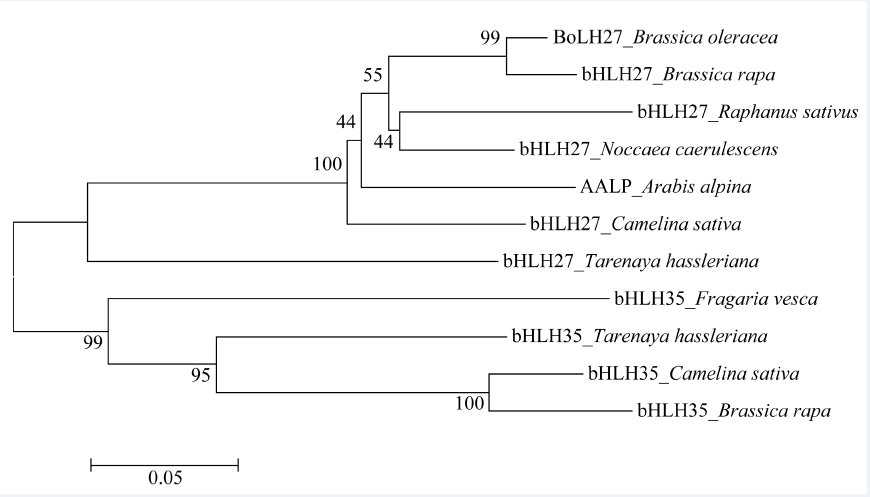
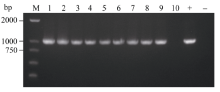
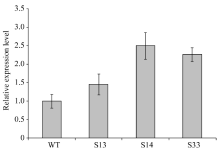
bHLH转录因子对植物生长发育、形态调控有重要作用, 为了研究BoLH27基因在甘蓝叶片发育及形态建成中的调控功能, 本文以甘蓝品种519为材料, 克隆出转录因子BoLH27基因。序列分析表明, 该基因含有一个795 bp的开放阅读框, 编码264个氨基酸, 具有一个保守的典型bHLH结构域。以农杆菌介导的遗传转化方法将正义BoLH27基因导入甘蓝品种519中以增强表达, 通过PCR筛选出9株T0代转基因单株, 结合T2代单株的qRT-PCR分析表明, 筛选的转基因植株内BoLH27基因的表达积累量明显高于野生型。试验基地隔离网内种植的转基因甘蓝表型明显, 主要表现为莲座期茎叶间距拉长、叶柄伸长以及植株茎和叶柄显示紫色, 植株叶片平展, 无向上向内卷曲趋势, 表明BoLH27基因可能对甘蓝叶片发育有重要的调控作用。
| [1] | Ludwig S R, Habera L F, Dellaporta S L, Wessler S R.Lc, a member of the maize R gene family responsible for tissue- specific anthocyanin production, encodes a protein similar to transcriptional activators and contains the myc-homology region. Proc Natl Acad Sci USA, 1989, 86: 7092-7096 |
| [2] | Ling H Q, Bauer P, Bereczky Z, Keller B, Ganal M.The tomato FER gene encoding a bHLH protein controls iron-uptake responses in roots.Proc Natl Acad Sci USA, 2002, 99: 13938-13943 |
| [3] | Kiribuchi K, Jikumaru Y, Kaku H, Minami E, Hasegawa M, Kodama O.Involvement of the basic helix-loop-helix transcription factor RERJ1 in wounding and drought stress responses in rice plants.Biosci Biotechnol Biochem, 2005, 69: 1042-1044 |
| [4] | Carreteropaulet L, Galstyan A, Roigvillanova I, Martínezgarcía J F, Bilbaocastro J R, Robertson D L.Genome-wide classification and evolutionary analysis of the bHLH family of transcription factors in Arabidopsis, poplar, rice, moss, and algae.Plant Physiol, 2010, 153: 1398-1412 |
| [5] | Pires N, Dolan L.Origin and diversification of basic-helix- loop-helix proteins in plants.Mol Biol Evol, 2010, 27: 862-874 |
| [6] | Grove C A, De M F, Barrasa M I, Newburger D E, Alkema M J, Bulyk M L.A multiparameter network reveals extensive divergence between C. elegans bHLH transcription factors. Cell, 2009, 138: 314-327 |
| [7] | Toledo-Ortiz G, Huq E, Quail P H.The Arabidopsis basic-helix-loop-helix transcription factor family. Plant Cell, 2003, 15: 1749-1770 |
| [8] | Li X, Duan X, Jiang H, Sun Y, Tang Y, Yuan Z.Genome-wide analysis of basic-helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Physiol, 2006, 141: 1167-1184 |
| [9] | Atchley W R, Terhalle W, Dress A.Positional dependence, cliques, and predictive motifs in the bHLH protein domain.J Mol Evol, 1999, 48: 501-516 |
| [10] | Heim M A, Jakoby M, Werber M, Martin C, Weisshaar B, Bailey P C.The basic helix-loop-helix transcription factor family in plants: a genome-wide study of protein structure and functional diversity.Mol Biol Evol, 2003, 20: 735-747 |
| [11] | Yin J, Chang X, Kasuga T, Mai B, Reid M S, Jiang C Z.A basic helix-loop-helix transcription factor, PhFBH4, regulates flower senescence by modulating ethylene biosynthesis pathway in petunia.Hortic Res, 2015, 2: 15059-15068 |
| [12] | Ko S S, Li M J, Sun-Ben K M, Ho Y C, Lin Y J, Chuang M H. The bHLH142 transcription factor coordinates with TDR1 to modulate the expression of EAT1 and regulate pollen development in rice.Plant Cell, 2014, 26: 2486-2504 |
| [13] | Takahashi Y, Ebisu Y, Kinoshita T, Doi M, Okuma E, Murata Y. bHLH transcription factors that facilitate K+ uptake during stomatal opening are repressed by abscisic acid through phosphorylation. Sci Signal, 2013, 6: ra48 |
| [14] | Shen Q, Lu X, Yan T, Fu X, Lv Z, Zhang F.The jasmonate-responsive AaMYC2 transcription factor positively regulates artemisinin biosynthesis in Artemisia annua. New Phytol, 2016, 210: 1269-1281 |
| [15] | Kurbidaeva A, Ezhova T, Novokreshchenova M.Arabidopsis thaliana ICE2 gene: phylogeny, structural evolution and functional diversification from ICE1. Plant Sci, 2014, 229: 10-22 |
| [16] | Makkena S, Lamb R S.The bHLH transcription factor SPATULA is a key regulator of organ size in Arabidopsis thaliana. Plant Signal Behav, 2013, 8: e24140 |
| [17] | An R, Liu X, Wang R, Wu H, Liang S, Shao J, Qi Y, An L, Yu F.The over-expression of two transcription factors, ABS5/bHLH30 and ABS7/MYB101, leads to upwardly curly leaves.PLoS One, 2014, 9: e107637 |
| [18] | Nath U, Crawford B C, Carpenter R, Coen E.Genetic control of surface curvature.Science, 2003, 299: 1404-1407 |
| [19] | Qin G, Gu H, Zhao Y, Ma Z, Shi G, Yang Y.An indole-3-acetic acid carboxyl methyltransferase regulates Arabidopsis leaf development.Plant Cell, 2005, 17: 2693-2704 |
| [20] | Sarvepalli K, Nath U.Hyper-activation of the TCP4 transcription factor in Arabidopsis thaliana accelerates multiple aspects of plant maturation. Plant J, 2011, 67: 595-607 |
| [21] | Efroni I, Blum E, Goldshmidt A, Eshed Y.A protracted and dynamic maturation schedule underlies Arabidopsis leaf development. Plant Cell, 2008, 20: 2293-2306 |
| [22] | 莫晓婷. 玉米逆境相关转录因子的克隆与初步分析. 中国农业科学院硕士学位论文. 北京, 2013 |
| Mo X T.Cloning and Functional Characterization of Stress-related Transcription Factors in Maize. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing,China, 2013 (in Chinese with English abstract) | |
| [23] | 何绍敏, 李春雨, 兰彩耘, 邹敏, 任雪松, 司军, 李成琼, 宋洪元. 转MLPK反义基因对甘蓝自交不亲和性的影响. 园艺学报, 2015, 42: 252-262 |
| He S M, Li C Y, Lan C Y, Zou M, Ren X S, Si J, Li C Q, Song H Y.Effect of antisense RNA of the MLPK gene on self-incompatibility in cabbage.Acta Hortic Sin, 2015, 42: 252-262 (in Chinese with English abstract) | |
| [24] | Heim M A, Jakoby M, Werber M, Martin C, Weisshaar B, Bailey P C.The basic helix-loop-helix transcription factor family in plants: a genome-wide study of protein structure and functional diversity.Mol Biol Evol, 2003, 20: 735-747 |
| [25] | Grotewold E, Sainz M B, Tagliani L, Hernandez J M, Bowen B, Chandler V L.Identification of the residues in the Myb domain of maize C1 that specify the interaction with the bHLH cofactor R.Proc Natl Acad Sci USA, 2000, 97: 13579-13584 |
| [26] | Bernhardt C, Lee M M, Gonzalez A, Zhang F, Lloyd A, Schiefelbein J.The bHLH genes GLABRA3 (GL3) and ENHANCER OF GLABRA3 (EGL3) specify epidermal cell fate in the Arabidopsis root. Development, 2003, 130: 6431-6439 |
| [27] | Zhang L Y, Bai M Y, Wu J, Zhu J Y, Wang H, Zhang Z.Antagonistic HLH/bHLH transcription factors mediate brassinosteroid regulation of cell elongation and plant development in rice and Arabidopsis. Plant Cell, 2009, 21: 3767-3780 |
| [28] | Meng Y, Li H, Wang Q, Liu B, Lin C.Blue light-dependent interaction between cryptochrome 2 and CIB1 regulates transcription and leaf senescence in soybean.Plant Cell, 2013, 25: 4405-4420 |
| [29] | Husbands A, Bell E M, Shuai B, Smith H M S, Springer P S. LATERAL ORGAN BOUNDARIES defines a new family of DNA-binding transcription factors and can interact with specific bHLH proteins.Nucl Acids Res, 2007, 35: 6663-6671 |
| [30] | Shuai B, Reynagapeña C G, Springer P S.The lateral organ boundaries gene defines a novel, plant-specific gene family.Plant Physiol, 2002, 129: 747-761 |
| [31] | Aguilar-Martínez J A, Sinha N. Analysis of the role of Arabidopsis class I TCP genes AtTCP7, AtTCP8, AtTCP22, and AtTCP23 in leaf development. Front Plant Sci, 2013, 8:e24140 |
| [32] | Guo Z, Fujioka S, Blancaflor E B, Miao S, Gou X, Li J.TCP1 modulates brassinosteroid biosynthesis by regulating the expression of the key biosynthetic gene DWARF4 in Arabidopsis thaliana. Plant Cell, 2010, 22: 1161-1173 |
| [33] | Palatnik J F, Allen E, Wu X, Schommer C, Schwab R, Carrington J C.Control of leaf morphogenesis by microRNAs.Nature, 2003, 425: 257-263 |
| [34] | Chandler V L, Radicella J P, Robbins T P, Chen J, Turks D.Two regulatory genes of the maize anthocyanin pathway are homologous: isolation of B utilizing R genomic sequences.Plant Cell, 1989, 1: 1175-1183 |
| [35] | de Vetten N, Quattrocchio F, Mol J, Koes R. The an11 locus controlling flower pigmentation in petunia encodes a novel WD-repeat protein conserved in yeast, plants, and animals.Genes Dev, 1997, 11: 1422-1434 |
| [36] | Baudry A, Caboche M, Lepiniec L.TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant Mol Biol, 2006, 46: 768-779 |
| [37] | Zhang F, Gonzalez A, Zhao M, Payne C T, Lloyd A.A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development, 2003, 130: 4859-4869 |
| [38] | Liu X F, Yin X R, Allan A C, Wang K L, Shi Y N, Huang Y J.The role of MrbHLH1, and MrMYB1, in regulating anthocyanin biosynthetic genes in tobacco and Chinese bayberry (Myrica rubra) during anthocyanin biosynthesis. Plant Cell Tissue Org, 2013, 115: 285-298 |
| [39] | Chiu L W, Zhou X, Burke S, Wu X, Prior R L, Li L.The purple cauliflower arises from activation of a MYB transcription factor.Plant Physiol, 2010, 154: 1470-1480 |
| [1] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [2] | 陈松余, 丁一娟, 孙峻溟, 黄登文, 杨楠, 代雨涵, 万华方, 钱伟. 甘蓝型油菜BnCNGC基因家族鉴定及其在核盘菌侵染和PEG处理下的表达特性分析[J]. 作物学报, 2022, 48(6): 1357-1371. |
| [3] | 秦璐, 韩配配, 常海滨, 顾炽明, 黄威, 李银水, 廖祥生, 谢立华, 廖星. 甘蓝型油菜耐低氮种质筛选及绿肥应用潜力评价[J]. 作物学报, 2022, 48(6): 1488-1501. |
| [4] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [5] | 张以忠, 曾文艺, 邓琳琼, 张贺翠, 刘倩莹, 左同鸿, 谢琴琴, 胡燈科, 袁崇墨, 廉小平, 朱利泉. 甘蓝S-位点基因SRK、SLG和SP11/SCR密码子偏好性分析[J]. 作物学报, 2022, 48(5): 1152-1168. |
| [6] | 王泽, 周钦阳, 刘聪, 穆悦, 郭威, 丁艳锋, 二宫正士. 基于无人机和地面图像的田间水稻冠层参数估测与评价[J]. 作物学报, 2022, 48(5): 1248-1261. |
| [7] | 周慧文, 丘立杭, 黄杏, 李强, 陈荣发, 范业赓, 罗含敏, 闫海锋, 翁梦苓, 周忠凤, 吴建明. 甘蔗赤霉素氧化酶基因ScGA20ox1的克隆及功能分析[J]. 作物学报, 2022, 48(4): 1017-1026. |
| [8] | 袁大双, 邓琬玉, 王珍, 彭茜, 张晓莉, 姚梦楠, 缪文杰, 朱冬鸣, 李加纳, 梁颖. 甘蓝型油菜BnMAPK2基因的克隆及功能分析[J]. 作物学报, 2022, 48(4): 840-850. |
| [9] | 黄成, 梁晓梅, 戴成, 文静, 易斌, 涂金星, 沈金雄, 傅廷栋, 马朝芝. 甘蓝型油菜BnAPs基因家族成员全基因组鉴定及分析[J]. 作物学报, 2022, 48(3): 597-607. |
| [10] | 王瑞, 陈雪, 郭青青, 周蓉, 陈蕾, 李加纳. 甘蓝型油菜白花基因InDel连锁标记开发[J]. 作物学报, 2022, 48(3): 759-769. |
| [11] | 谢琴琴, 左同鸿, 胡燈科, 刘倩莹, 张以忠, 张贺翠, 曾文艺, 袁崇墨, 朱利泉. 甘蓝自交不亲和相关基因BoPUB9的克隆及表达分析[J]. 作物学报, 2022, 48(1): 108-120. |
| [12] | 王渭霞, 赖凤香, 胡海燕, 何佳春, 魏琪, 万品俊, 傅强. 超低温11年保存期对转基因作物基体标准样品核酸检测的影响[J]. 作物学报, 2022, 48(1): 238-248. |
| [13] | 王艳花, 刘景森, 李加纳. 整合GWAS和WGCNA筛选鉴定甘蓝型油菜生物产量候选基因[J]. 作物学报, 2021, 47(8): 1491-1510. |
| [14] | 左香君, 房朋朋, 李加纳, 钱伟, 梅家琴. 有毛野生甘蓝(Brassica incana)抗蚜虫特性研究[J]. 作物学报, 2021, 47(6): 1109-1113. |
| [15] | 李杰华, 端群, 史明涛, 吴潞梅, 柳寒, 林拥军, 吴高兵, 范楚川, 周永明. 新型抗广谱性除草剂草甘膦转基因油菜的创制及其鉴定[J]. 作物学报, 2021, 47(5): 789-798. |
|
||