欢迎访问作物学报,今天是
作物学报 ›› 2019, Vol. 45 ›› Issue (3): 460-468.doi: 10.3724/SP.J.1006.2019.84002
何宁1,王雪扬1,曹良子1,曹大为1,洛育1,姜连子2,孟英1,冷春旭1,唐晓东1,李一丹1,万书明1,卢环1,程须珍3,*( )
)
Ning HE1,Xue-Yang WANG1,Liang-Zi CAO1,Da-Wei CAO1,Yu LUO1,Lian-Zi JIANG2,Ying MENG1,Chun-Xu LENG1,Xiao-Dong TANG1,Yi-Dan LI1,Shu-Ming WAN1,Huan LU1,Xu-Zhen CHENG3,*()
摘要:
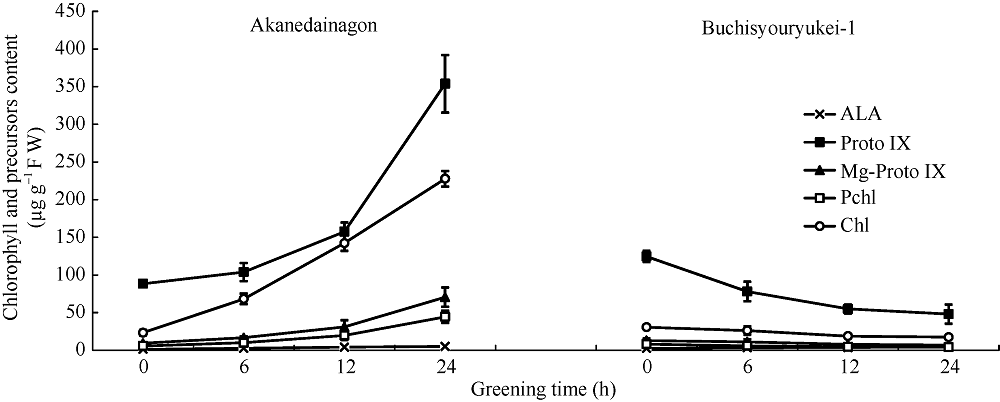
探讨不同低温和光照条件下小豆苗期的冷害发生机制及引起初生叶黄化和叶绿素合成的受阻位点, 旨在为小豆耐寒新品种选育及栽培提供理论依据。选择2个对温度和光照反应不同的日本小豆品种, 利用人工气候室再现小豆苗期的低温障碍, 研究低温遮光处理(昼夜10~13°C, 2%遮光) 18 d和28 d对小豆苗期H2O2、SOD、CAT、APX、叶绿素的影响; 利用植物生长箱再现小豆苗期的黄化障碍, 研究不同的低温处理长度(1 d、3 d、5 d、7 d; 10°C, 50 μmol m -2s -1)和暗处理长度(25°C, 黑暗1 d、3 d、5 d、7 d)对绿化后(24 h、25°C、62.5 μmol m -2s -1)叶绿素合成能力及受阻位点的影响。苗期小豆耐低温和不耐低温品种的最大差异是长期低温遮光处理的H2O2含量和SOD活性。长期低温遮光处理后不耐低温品种的H2O2含量是耐低温品种的约66倍, 但随着绿化处理时间的延长, 抗氧化酶活性和叶绿素含量急剧下降直至8 h后消失。与低温处理相比, 暗处理才是造成叶绿素合成能力差异的主要原因。对叶绿素合成中间产物的研究表明, 从ALA向Proto IX的转化可能受阻, 最终导致叶绿素合成受阻, 叶绿素含量下降。说明H2O2含量和SOD活性可能与小豆苗期耐冷性关系更密切。引起小豆苗期叶绿素合成受阻位点是Proto IX的转化。
| [1] | 徐宁, 程须珍, 王丽侠, 王素华, 刘长友, 孙蕾, 梅丽 . 用于中国小豆种质资源遗传多样性分析SSR分子标记筛选及应用. 作物学报, 2009,35:219-227. |
| Xu N, Cheng X Z, Wang L X, Wang S H, Liu C Y, Sun L, Mei L . Screening and application of SSR molecular markers for genetic diversity analysis of Chinese adzuki bean germplasm resources. Acta Agron Sin, 2009,35:219-227 (in Chinese with English abstract). | |
| [2] | 村田吉平 . 北海道における作物育種. 札幌: 北海道協同組合通信社, 1998. pp 139-155. |
| Murata K P. Hokkaido Plant Breeding. Sapporo: Hokkaido Agricultural Cooperatives Publisher, 1998. pp 139-155 (in Japanese). | |
| [3] | 新免輝夫 . 現代植物生理学. 东京: 環境応答, 1991. pp 142-158. |
| Shinme T. Modern Plant Physiology. Tokyo: Institute of Society Press, 1991. pp 142-158(in Japanese). | |
| [4] | Northen R T . Home Orchid Growing, 3rd edn. New York: Van Nostrand Reinhold, 1970. pp 19-28. |
| [5] | 横田明穂 . 植物ストレスにおける応答. 東京: 学会出版センター, 2002. pp 209-224. |
| Yokota M H. Introduction to Plant Molecular Physiology. Tokyo: Institute of Society Press, 2002. pp 209-224(in Japanese). | |
| [6] | 李進才 , 趙習コウ, 松井鋳一郎 . 光ストレスおよび遮光栽培におけるCattleyaとCymbidium葉の抗酸化酵素活性および色素含量の変化. 園学雑, 2001,70:372-379. |
| Li J C, Zhao X K, Matsui I . Effect of light stress and shading cultivation on antioxidant enzyme activity and pigment content of cattleya and cymbidium leaves. Jpn Hortic J, 2001,70:372-379 (in Japanese). | |
| [7] |
Salin W L . Toxic oxygen species and protective systems of the chloroplast. Physiol Plant, 1988,72:681-689.
doi: 10.1111/ppl.1988.72.issue-3 |
| [8] | Shen W K, Nada S, Tachibana S . Oxygen radical generation in chilled leaves of cucumber ( Cucumis sativus L.) cultivars with different tolerances to chilling temperatures. Engei Gakkai Zasshi, 2008,68:780-787. |
| [9] |
Erich W, Laties G G . Quantification of hydrogen peroxide in plant extracts by the chemiluminescence reaction with luminal. Hytochemistry, 1982,21:827-831.
doi: 10.1016/0031-9422(82)80073-3 |
| [10] | 沈利星 . 雑草科学実験法. 第5項植物の抗酸化活性測定法. 東京: 日本雑草学会発行, 2001. pp 296-298. |
| Shen L X . Methods of Scientific Experiment of Weeds and Determination of Antioxidant Activity of Plants. Tokyo: The Japanese Weed Society Publishers, 2001. pp 296-298 (in Japanese). | |
| [11] | 加藤荣 . 光合成研究法. 东京: 日本共立出版社, 1981. pp 40-41. |
| Katou E. Research Methods of Photosynthesis. Tokyo: Japan Co-publisher, 1981. pp 40-41(in Japanese). | |
| [12] | 何寧, 小嶋道之, 黒澤聪, 加藤清明 . 低温遮光処理アズキ初生葉の過酸化水素含量と抗酸化酵素活性に及ぼす影響. 日本作物学会記事, 2006,75:360-365. |
| He N, Kojima M Y, Kurosawa A, Kato K . Effects of chilling and shading on hydrogen peroxide content and activity of oxidation-inhibiting enzymes in the primary leaves of adzuki beans. Jpn J Crop Sci, 2006,75:360-365 (in Japanese). | |
| [13] |
Harpaz-Saad S, Azoulay T, Arazi T, Ben-Yaakov E, Mett A, Hörtensteiner S, Gidoni D, Gal-On A, Goldschmidt E E, Eyal Y . Chlorophyllase is a rate-limiting enzyme in chlorophyll catabolism and is posttranslationally regulated. Plant Cell, 2007,19:1007-1022.
doi: 10.1105/tpc.107.050633 |
| [14] | Von Gromoff E D, Alawady A, Meinecke L, Grimm B, Beck C F . Heme, a plastid-derived regulator of nuclear gene expression in chlamydomonas. Plant Cell, 2008,20:552-567. |
| [15] |
王平荣, 张帆涛, 高家旭, 孙小秋, 邓晓建 . 高等植物叶绿素生物合成的研究进展. 西北植物学报, 2009,29:629-636.
doi: 10.3321/j.issn:1000-4025.2009.03.032 |
|
Wang P R, Zhang F T, Gao J X, Sun X Q, Deng X J . An overview of chlorophyll biosynthesis in higher plants. Acta Bot Boreali- Occident Sin, 2009,29:629-636 (in Chinese with English abstract).
doi: 10.3321/j.issn:1000-4025.2009.03.032 |
|
| [16] |
Okuda T Y, Matuda A, Yamanaka S, Sagisaka S . Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol, 1991,97:1265-1267.
doi: 10.1104/pp.97.3.1265 pmid: 16668520 |
| [17] |
Fridovich I . Biological effects of the superoxide radicals. Arch Biochem Biophys, 1986,247:1-11.
doi: 10.1016/0003-9861(86)90526-6 pmid: 3010872 |
| [18] | 重岡成 . 活性酸素代謝の応答機構. 光合成生物の活性酸素代謝の応答機構. 生物工学会誌, 2001,79:303-322. |
| Shigeoka N R . The response mechanism of photosynthetic biological reactive oxygen metabolism. Jpn J Bioengineer, 2001,79:303-322 (in Japanese). | |
| [19] |
Azevedo R A, Alas R M, Smith R J, Lea P J . Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiol Plant, 1998,104:280-292.
doi: 10.1034/j.1399-3054.1998.1040217.x |
| [20] |
Foyer C H, Mullineaux P . Causes of photooxidative stress and amelioration of defense systems in plants. Acta Ophthalmol, 1994,44:276-315.
doi: 10.1111/j.1755-3768.1966.tb08068.x |
| [21] | 宮尾光恵, 水澤直樹 . 強光環境から身を守る植物の防御機構. 化学と生物, 1999,37:396-400. |
| Miyao H E, Mizusawa N K . The plant defense mechanism obtained from the strong light environment. Jpn Chem Biol, 1999,37:396-400 (in Japanese). | |
| [22] |
王宝增 . 叶绿素降解代谢的研究进展. 生物学教学, 2010,35(2):7-9.
doi: 10.3969/j.issn.1004-7549.2010.02.004 |
|
Wang B Z . Research progress of chlorophyll degradation metabolism. Biol Teach, 2010,35(2):7-9 (in Chinese with English abstract).
doi: 10.3969/j.issn.1004-7549.2010.02.004 |
|
| [23] |
Gopal K, Pattanayak G K, Biswal A K, Reddy V S, Tripathy B C . Light-dependent regulation of chlorophyll b biosynthesis in chlorophyllide a oxygenase overexpressing tobacco plants. Biochem Biophys Res Commun, 2005,326:466-471.
doi: 10.1016/j.bbrc.2004.11.049 pmid: 15582600 |
| [24] | 田中歩, 平島真澄, 田中亮一 . クロロフェル代謝と植物の生育. 化学と生物, 2004,42:2-26. |
| Tanaka A, Hirasima M S, Tanaka L Y . The development of chlorophyll metabolism and plant. Chem Biol, 2004,42:2-26 (in Japanese with English abstract). |
| [1] | 李增强, 丁鑫超, 卢海, 胡亚丽, 岳娇, 黄震, 莫良玉, 陈立, 陈涛, 陈鹏. 铅胁迫下红麻生理特性及DNA甲基化分析[J]. 作物学报, 2021, 47(6): 1031-1042. |
| [2] | 项洪涛, 李琬, 郑殿峰, 王诗雅, 何宁, 王曼力, 杨纯杰. 幼苗期淹水胁迫及喷施烯效唑对小豆生理和产量的影响[J]. 作物学报, 2021, 47(3): 494-506. |
| [3] | 田文刚,朱雪峰,宋雯,程文翰,薛飞,朱华国. 异源表达棉花S-腺苷甲硫氨酸脱羧酶(GhSAMDC1)基因提高了拟南芥抗盐能力[J]. 作物学报, 2019, 45(7): 1017-1028. |
| [4] | 万丽丽, 王转茸, 辛强, 董发明, 洪登峰, 杨光圣. BnA7HSP70分子伴侣结合蛋白超表达能够提高甘蓝型油菜耐旱性[J]. 作物学报, 2018, 44(04): 483-492. |
| [5] | 马燕明,陈春海,杨凯,李奕松,赵波,李江,李永强,万平. 野生小豆和栽培小豆microRNA全基因组鉴定与比较分析[J]. 作物学报, 2016, 42(09): 1273-1281. |
| [6] | 卢霖,董志强*,董学瑞,李光彦. 乙矮合剂对不同密度夏玉米花粒期不同部位叶片衰老特性的影响[J]. 作物学报, 2016, 42(04): 561-573. |
| [7] | 徐金刚,吕川根,刘莉,吕春芳,马静,夏士健,陈国祥,高志萍. 水稻光氧化突变体812HS的光合和抗氧化特性[J]. 作物学报, 2016, 42(04): 574-582. |
| [8] | 张英华,杨佑明,曹莲,郝杨凡,黄菁,李金鹏,姚得秀,王志敏*. 灌浆期高温对小麦旗叶与非叶器官光合和抗氧化酶活性的影响[J]. 作物学报, 2015, 41(01): 136-144. |
| [9] | 白鹏,程须珍*,王丽侠,王素华,陈红霖. 小豆遗传差异、群体结构和连锁不平衡水平的SSR分析[J]. 作物学报, 2014, 40(05): 788-797. |
| [10] | 丁秀文,张国良,戴其根,朱青. 1,2,4-三氯苯胁迫对水稻分蘖盛期植株生长和生理特性的影响[J]. 作物学报, 2014, 40(03): 487-496. |
| [11] | 刘长友,范保杰,曹志敏,苏秋竹,王彦,张志肖,程须珍,田静. 利用SSR标记分析野生小豆及其近缘野生植物的遗传多样性[J]. 作物学报, 2014, 40(01): 174-180. |
| [12] | 王永军,杨今胜,袁翠平,柳京国,李登海,董树亭. 超高产夏玉米花粒期不同部位叶片衰老与抗氧化酶特性[J]. 作物学报, 2013, 39(12): 2183-2191. |
| [13] | 张会慧,田褀,刘关君,胡彦波,吴翔宇,田野,李鑫,孙广玉. 转2-Cys Prx基因烟草抗氧化酶和PSII电子传递对盐和光胁迫的响应[J]. 作物学报, 2013, 39(11): 2023-2029. |
| [14] | 侯鹏飞,马俊青,赵鹏飞,张欢玲,赵会杰,刘华山,赵一丹,汪月霞. 外源甜菜碱对干旱胁迫下小麦幼苗叶绿体抗氧化酶及psbA基因表达的调节[J]. 作物学报, 2013, 39(07): 1319-1324. |
| [15] | 杨东清,王振林*,尹燕枰,倪英丽,杨卫兵,蔡铁,彭佃亮,徐彩龙,崔正勇,刘铁宁,徐海成. 外源ABA和6-BA对不同持绿型小麦旗叶衰老的影响及其生理机制[J]. 作物学报, 2013, 39(06): 1096-1104. |
|
||