欢迎访问作物学报,今天是
作物学报 ›› 2019, Vol. 45 ›› Issue (11): 1615-1627.doi: 10.3724/SP.J.1006.2019.91009
• 作物遗传育种·种质资源·分子遗传学 • 下一篇
张宏娟1,2,李玉莹2,3,苗丽丽2,王景一2,李超男2,杨德龙1,*( ),毛新国1,2,*(),景蕊莲2
),毛新国1,2,*(),景蕊莲2
ZHANG Hong-Juan1,2,LI Yu-Ying2,3,MIAO Li-Li2,WANG Jing-Yi2,LI Chao-Nan2,YANG De-Long1,*(),MAO Xin-Guo1,2,*(),JING Rui-Lian2
摘要:
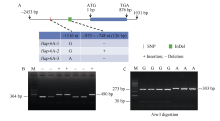
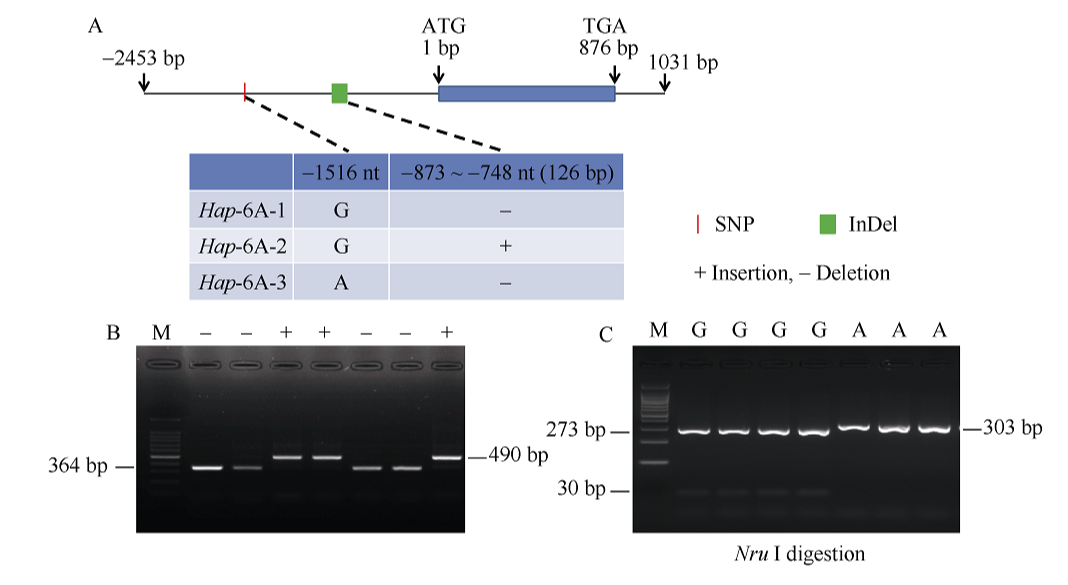
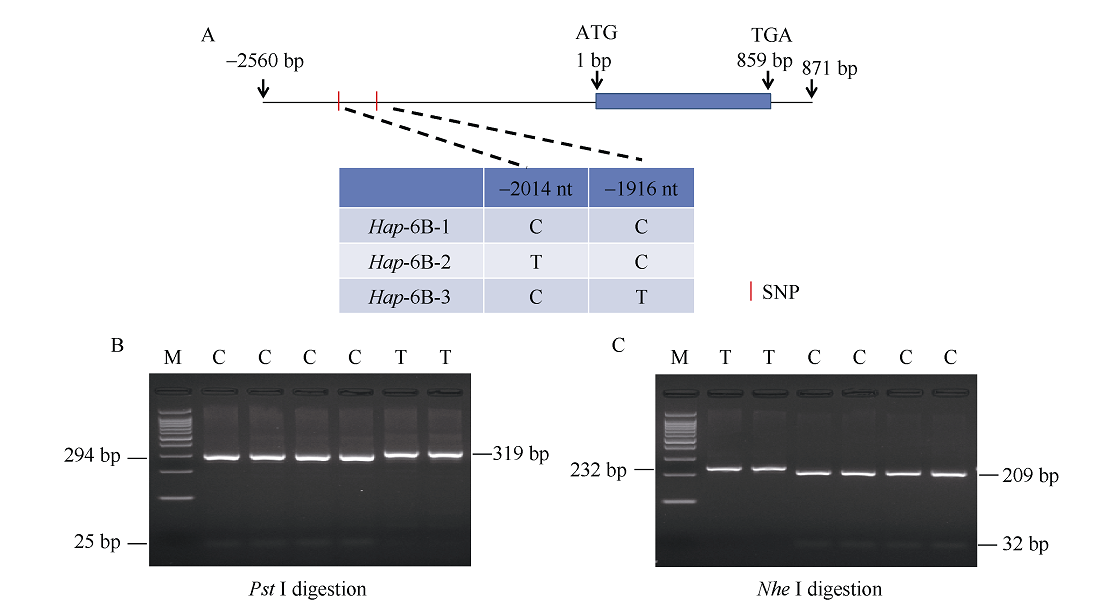
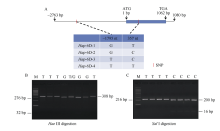
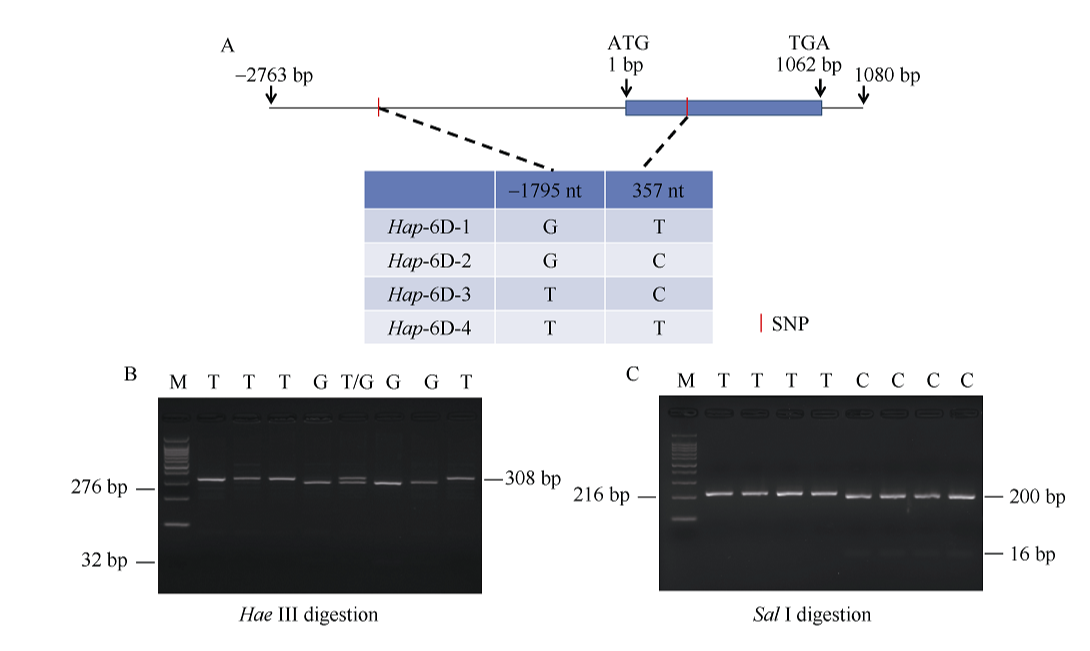
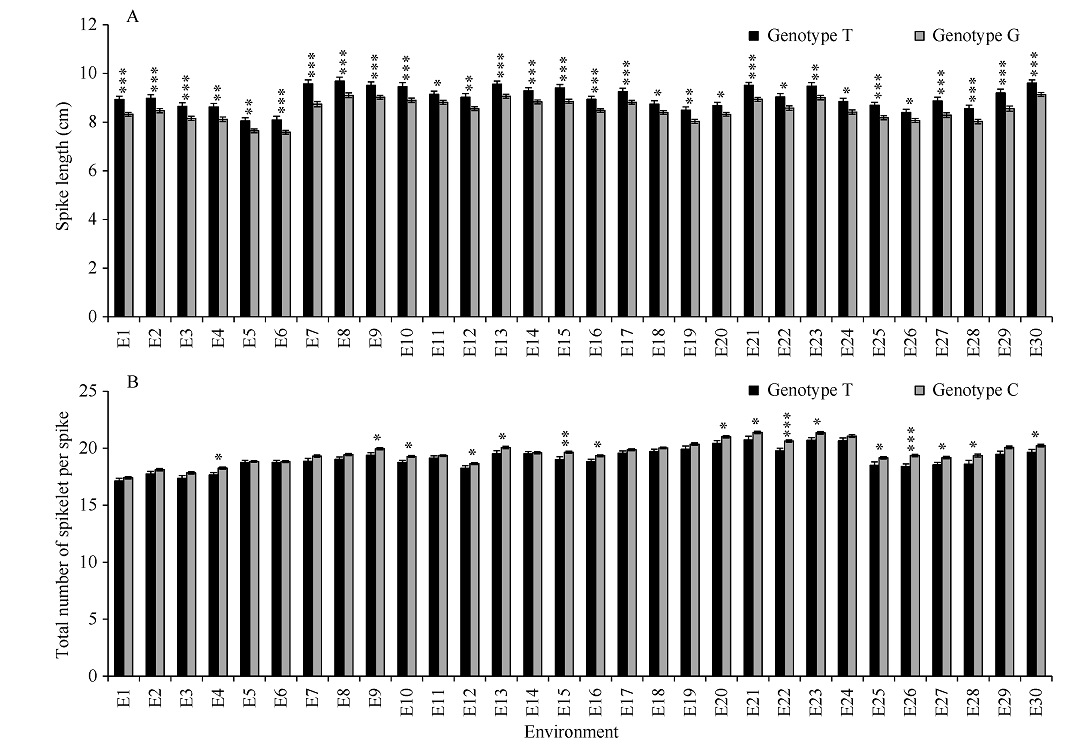
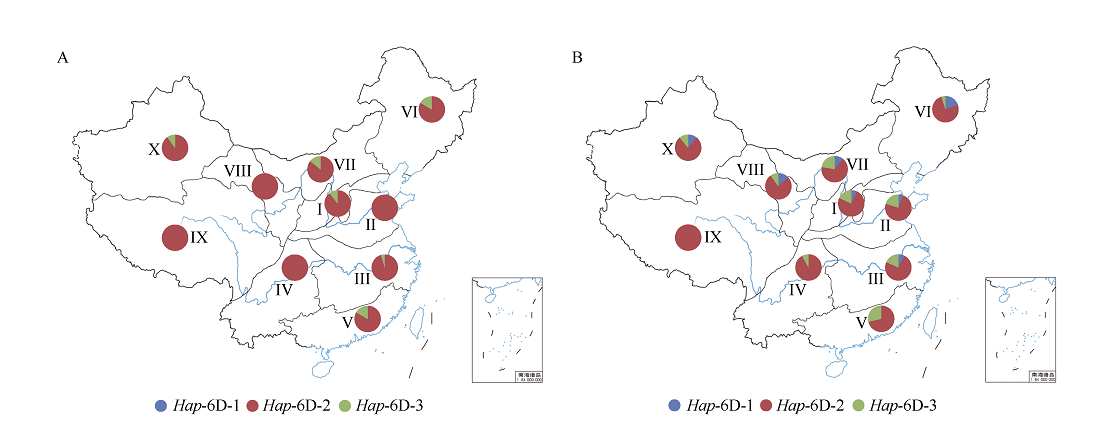
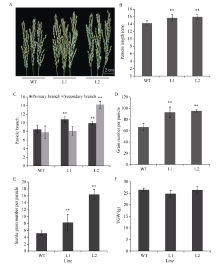
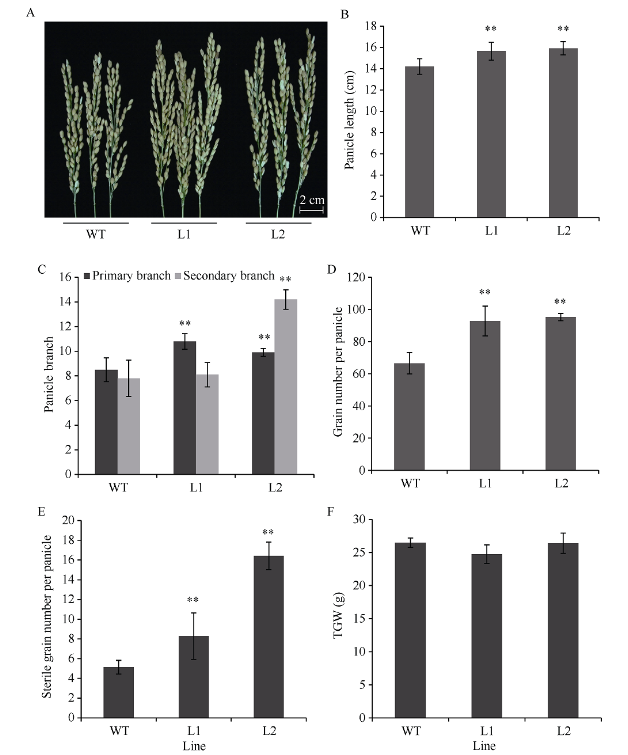
NAC转录因子是植物特有的一类转录因子, 在植物生长发育和逆境胁迫应答反应中发挥着重要作用。前期研究表明, TaNAC67参与对多种逆境胁迫的应答, 过量表达能增强拟南芥的抗逆性。为进一步揭示其在调控小麦主要农艺性状发育方面的作用, 本研究以36份普通小麦组成的高多态性群体为材料, 测序分析了TaNAC67-6A、TaNAC67-6B、TaNAC67-6D序列多态性, 发现TaNAC67-6A启动子区-1516 nt有1个A/G转换SNP, 在-873~ -748 nt处有1个126 bp的InDel; TaNAC67-6B启动子区-2014和-1916 nt处各有1个C/T转换SNP; TaNAC67-6D启动子区-1795 nt有1个T/G颠换SNP, 编码区357 nt处有1个C/T转换SNP。根据多态性分别开发了功能分子标记, 扫描由282份普通小麦构成的自然群体, 并将基因型和表型性状检测结果进行关联分析, TaNAC67-6A、TaNAC67-6B的标记与表型性状无显著关联, 而TaNAC67-6D的2个标记SNP-D-1和SNP-D-2分别与小麦穗长和每穗小穗数显著相关。单倍型分析发现, 自然群体中存在3种主要单倍型, 其中Hap-6D-3是增加穗长和每穗小穗数的最优单倍型, 在我国小麦育种历史中受到了正向选择。转基因水稻表型分析发现, TaNAC67过表达能显著增加水稻主穗穗长、穗分枝和穗粒数, 验证了小麦关联分析结果。因此, TaNAC67-6D可用于改良农作物穗部性状, 其分子标记可用于小麦分子标记辅助选择育种。
| [1] | Huang Q, Wang Y, Li B, Chang J, Chen M, Li K, Yang G, He G . TaNAC29, a NAC transcription factor from wheat, enhances salt and drought tolerance in transgenic Arabidopsis. BMC Plant Biol, 2015,15:268. |
| [2] | Jin C, Li K Q, Xu X Y, Zhang H P, Chen H X, Chen Y H, Hao J, Wang Y, Huang X S, Zhang S L . A novel NAC transcription factor, PbeNAC1, of Pyrus betulifolia confers cold and drought tolerance via interacting with PbeDREBs and activating the expression of stress-responsive genes. Front Plant Sci, 2017,8:1049. |
| [3] | Hu H, You J, Fang Y, Zhu X, Qi Z, Xiong L . Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol Biol, 2010,72:567-568. |
| [4] | Aida M, Ishida T, Fukaki H, Fujisawa H, Tasaka M . Genes involved in organ separation in Arabidopsis: an analysis of the cup-shaped cotyledon mutant. Plant Cell, 1997,9:841-857. |
| [5] | Taoka K, Yanagimoto Y, Daimon Y, Hibara K, Aida M, Tasaka M . The NAC domain mediates functional specificity of cup-shaped cotyledon proteins. Plant J, 2004,40:462. |
| [6] | Nakashima K, Tran L S, Van Nguyen D, Fujita M, Maruyama K, Todaka D, Ito Y, Hayashi N, Shinozaki K, Yamaguchi-Shinozaki K . Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J, 2007,51:617-630. |
| [7] | Nuruzzaman M, Manimekalai R, Sharoni A M, Satoh K, Kondoh H, Ooka H, Kikuchi S . Genome-wide analysis of NAC transcription factor family in rice. Gene, 2010,465:30-44. |
| [8] | Souer E, van Houwelingen Adèle, Kloos D, Mol J, Koes R . Kloos The no apical meristem gene of Petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell, 1996,85:159-170. |
| [9] | Chen X, Cheng J, Chen L, Zhang G, Huang H, Zhang Y, Xu L . Auxin-independent NAC pathway acts in response to explant- specific wounding and promotes root tip emergence during de novo root organogenesis in Arabidopsis. Plant Physiol, 2016,170:2136-2145. |
| [10] | Redillas M C, Jeong J S, Kim Y S, Jung H, Bang S W, Choi Y D, Ha S H, Reuzeau C, Kim J K . The overexpression of OsNAC9 alters the root architecture of rice plants enhancing drought resistance and grain yield under field conditions. Plant Biotechnol J, 2012,10:792-805. |
| [11] | Shim J S, Oh N, Chung P J, Kim Y S, Choi Y D, Kim J K . Overexpression of OsNAC14 improves drought tolerance in rice. Front Plant Sci, 2018,9:310. |
| [12] | Lee D K, Chung P J, Jeong J S, Jang G, Bang S W, Jung H, Kim Y S, Ha S H, Choi Y D, Kim J K . The rice OsNAC6 transcription factor orchestrates multiple molecular mechanisms involving root structural adaptions and nicotianamine biosynthesis for drought tolerance. Plant Biotechnol J, 2017,15:754-764. |
| [13] | Li J, Guo G H, Guo W W, Guo G G, Tong D, Ni Z F, Sun Q X, Yao Y Y . miRNA164-directed cleavage of ZmNAC1 confers lateral root development in maize (Zea mays L.). BMC Plant Biol, 2012,12:220. |
| [14] | Chen D D, Chai S C, Mcintyre C L, Xue G P . Overexpression of a predominantly root-expressed NAC transcription factor in wheat roots enhances root length, biomass and drought tolerance. Plant Cell Rep, 2018,37:225-237. |
| [15] | Chen D, Richardson T, Chai S, Lynne Mcintyre C, Rae A L, Xue G P . Drought-up-regulated TaNAC69-1 is a transcriptional repressor of TaSHY2 and TaIAA7, and enhances root length and biomass in wheat. Plant Cell Physiol, 2016,57:2076-2090. |
| [16] | Li W, Li X X, Chao J T, Zhang Z L, Wang W F, Guo Y F . NAC family transcription factors in tobacco and their potential role in regulating leaf senescence. Front Plant Sci, 2018,9:1900 |
| [17] | Zhao F L, Ma J H, Li L B, Fan S L, Guo Y N, Song M Z, Wei H L, Pang C Y, Yu S X . GhNAC12, a neutral candidate gene, leads to early aging in cotton(Gossypium hirsutum L.). Gene, 2015,576:268-274. |
| [18] | Ren T T, Wang J W, Zhao M M, Gong X M, Wang S X, Wang G, Zhou C J . Involvement of NAC transcription factor SiNAC1 in a positive feedback loop via ABA biosynthesis and leaf senescence in foxtail millet. Planta, 2017,247:1-16. |
| [19] | El Mannai Y, Akabane K, Hiratsu K, Satoh-Nagasawa N, Wabiko H . The NAC transcription factor gene OsY37 (ONAC011) promotes leaf senescence and accelerates heading time in rice. Int J Mol Sci, 2017,18:2165. |
| [20] | Christiansen M W, Matthewman C, Podzimska-Sroka D, O’Shea C, Lindemose S, Mollegaard N E, Holme I B, Hebelstrup K, Skriver K, Gregersen P L . Barley plants over-expressing the NAC transcription factor gene HvNAC005 show stunting and delay in development combined with early senescence. J Exp Bot, 2016,67:5259-5273. |
| [21] | Collinge M, Boller T . Differential induction of two potato genes, Stprx2 and StNAC, in response to infection by Phytophthora infestans and to wounding. Plant Mol Biol, 2001,46:521-529. |
| [22] | Xia N, Zhang G, Liu X Y, Deng L, Cai G L, Zhang Y, Wang X J, Zhao J, Huang L L, Kang Z S . Characterization of a novel wheat NAC transcription factor gene involved in defense response against stripe rust pathogen infection and abiotic stresses. Mol Biol Rep, 2010,37:3703-3712. |
| [23] | Wang B, Wei J, Song N, Wang N, Zhao J, Kang Z S . A novel wheat NAC transcription factor, TaNAC30, negatively regulates resistance of wheat to stripe rust. J Integr Plant Biol, 2018,60:432-443. |
| [24] | Wang Z, Xia Y, Lin S, Wang Y, Guo B, Song X, Ding S, Zheng L, Feng R, Chen S, Bao Y, Sheng C, Zhang X, Wu J, Niu D, Jin H, Zhao H . Osa-miR164a targets OsNAC60 and negatively regulates rice immunity against the blast fungus Magnaporthe oryzae. Plant J, 2018,95:584-597. |
| [25] | Liu Q, Yan S J, Huang W J, Yang J Y, Dong J F, Zhang S H, Zhao J L, Yang T F, Mao X X, Zhu X Y . NAC transcription factor ONAC066 positively regulates disease resistance by suppressing the ABA signaling pathway in rice. Plant Mol Biol, 2018,98:289-302. |
| [26] | Mao C, Ding J, Zhang B, Xi D, Ming F . OsNAC2 positively affects salt-induced cell death and binds to the OsAP37 and OsCOX11 promoters. Plant J, 2018,94:454-468. |
| [27] | Shen J, Lü B, Luo L, He J, Mao C, Xi D, Ming F . The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci Rep, 2017,7:40641. |
| [28] | Chung P J, Jung H, Choi Y D, Kim J K . Genome-wide analyses of direct target genes of four rice NAC-domain transcription factors involved in drought tolerance. BMC Genom, 2018,19:40. |
| [29] | 卢敏, 张登峰, 石云素, 宋燕春, 黎裕, 王天宇 . 玉米胁迫诱导表达基因ZmSNAC1的功能分析. 作物学报, 2013,39:2177-2182. |
| Lu M, Zhang D F, Shi Y S, Song Y C, Li Y, Wang T Y . Overexpression of a stress induced maize NAC transcription factor gene, ZmSNAC1, improved drought and salt tolerance in Arabidopsis. Acta Agron Sin, 2013,39:2177-2182 (in Chinese with English abstract). | |
| [30] | Mao H, Yu L, Han R, Li Z, Liu H . ZmNAC55, a maize stress-responsive NAC transcription factor, confers drought resistance in transgenic Arabidopsis. Plant Physiol Biochem, 2016,105:55-66. |
| [31] | Xia N, Zhang G, Yan F, Zhu L, Xu L S, Chen X M, Liu B O . TaNAC8, a novel NAC transcription factor gene in wheat, responds to stripe rust pathogen infection and abiotic stresses. Physiol Mol Plant Pathol, 2010,74:394-402. |
| [32] | Mao X G, Zhang H Y, Qian X Y, Li A, Zhao G Y, Jing R L . TaNAC2, a NAC-type wheat transcription factor conferring enhanced multiple abiotic stress tolerances in Arabidopsis. J Exp Bot, 2012,63:2933-2946. |
| [33] | Mao X G, Chen S S, Li A, Zhai C C, Jing R L . Novel NAC transcription factor TaNAC67 confers enhanced multi-abiotic stress tolerances in Arabidopsis. PLoS One, 2014,9:e84359. |
| [34] | 金善宝 . 中国小麦学. 北京: 中国农业出版社, 1996. pp 95-124. |
| Jin S B . Chinese Wheat. Beijing: China Agriculture Press, 1996. pp 95-124(in Chinese). | |
| [35] | Shiriga K, Sharma R, Kumar K, Yadav S K, Hossain F, Thirunavukkarasu N . Genome-wide identification and expression pattern of drought-responsive members of the NAC family in maize. Meta Gene, 2014,2:407-417. |
| [36] | Singh A K, Sharma V, Pal A K, Acharya V, Ahuja P S . Genome-wide organization and expression profiling of the NAC transcription factor family in potato (italic>Solanum tuberosum.). DNA Res, 2013,20:403-423. |
| [37] | Borrill P, Harrington S A, Uauy C . Genome-wide sequence and expression analysis of the NAC transcription factor family in polyploid wheat. G3: Genes Genom Genet, 2017,7:3019-3029. |
| [38] | Huysmans M, Buono R A, Skorzinski N, Radio M C, De Winter F, Parizot B, Mertens J, Karimi M, Fendrych M, Nowack M K . NAC transcription factors ANAC087 and ANAC046 control distinct aspects of programmed cell death in the Arabidopsis columella and lateral root cap. Plant Cell, 2018,30:2197-2213. |
| [39] | Guo S, Dai S, Singh P K, Wang H, Wang Y, Tan J L H, Wee W, Ito T . A membrane-bound NAC-like transcription factor OsNTL5 represses the flowering in Oryza sativa. Front Plant Sci, 2018,9:555. |
| [1] | 陈玲玲, 李战, 刘亭萱, 谷勇哲, 宋健, 王俊, 邱丽娟. 基于783份大豆种质资源的叶柄夹角全基因组关联分析[J]. 作物学报, 2022, 48(6): 1333-1345. |
| [2] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [3] | 郭星宇, 刘朋召, 王瑞, 王小利, 李军. 旱地冬小麦产量、氮肥利用率及土壤氮素平衡对降水年型与施氮量的响应[J]. 作物学报, 2022, 48(5): 1262-1272. |
| [4] | 孙思敏, 韩贝, 陈林, 孙伟男, 张献龙, 杨细燕. 棉花苗期根系分型及根系性状的关联分析[J]. 作物学报, 2022, 48(5): 1081-1090. |
| [5] | 朱峥, 王田幸子, 陈悦, 刘玉晴, 燕高伟, 徐珊, 马金姣, 窦世娟, 李莉云, 刘国振. 水稻转录因子WRKY68在Xa21介导的抗白叶枯病反应中发挥正调控作用[J]. 作物学报, 2022, 48(5): 1129-1140. |
| [6] | 陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展[J]. 作物学报, 2022, 48(4): 781-790. |
| [7] | 刘丹, 周彩娥, 王晓婷, 吴启蒙, 张旭, 王琪琳, 曾庆东, 康振生, 韩德俊, 吴建辉. 利用集群分离分析结合高密度芯片快速定位小麦成株期抗条锈病基因YrC271[J]. 作物学报, 2022, 48(3): 553-564. |
| [8] | 付美玉, 熊宏春, 周春云, 郭会君, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 徐延浩, 刘录祥. 小麦矮秆突变体je0098的遗传分析与其矮秆基因定位[J]. 作物学报, 2022, 48(3): 580-589. |
| [9] | 冯健超, 许倍铭, 江薛丽, 胡海洲, 马英, 王晨阳, 王永华, 马冬云. 小麦籽粒不同层次酚类物质与抗氧化活性差异及氮肥调控效应[J]. 作物学报, 2022, 48(3): 704-715. |
| [10] | 刘运景, 郑飞娜, 张秀, 初金鹏, 于海涛, 代兴龙, 贺明荣. 宽幅播种对强筋小麦籽粒产量、品质和氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 716-725. |
| [11] | 马红勃, 刘东涛, 冯国华, 王静, 朱雪成, 张会云, 刘静, 刘立伟, 易媛. 黄淮麦区Fhb1基因的育种应用[J]. 作物学报, 2022, 48(3): 747-758. |
| [12] | 黄莉, 陈玉宁, 罗怀勇, 周小静, 刘念, 陈伟刚, 雷永, 廖伯寿, 姜慧芳. 花生种子大小相关性状QTL定位研究进展[J]. 作物学报, 2022, 48(2): 280-291. |
| [13] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [14] | 王洋洋, 贺利, 任德超, 段剑钊, 胡新, 刘万代, 郭天财, 王永华, 冯伟. 基于主成分-聚类分析的不同水分冬小麦晚霜冻害评价[J]. 作物学报, 2022, 48(2): 448-462. |
| [15] | 陈新宜, 宋宇航, 张孟寒, 李小艳, 李华, 汪月霞, 齐学礼. 干旱对不同品种小麦幼苗的生理生化胁迫以及外源5-氨基乙酰丙酸的缓解作用[J]. 作物学报, 2022, 48(2): 478-487. |
|
||