欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (12): 1831-1838.doi: 10.3724/SP.J.1006.2020.03017
宋欣冉1( ), 胡书婷2(), 张凯2, 崔则瑾2, 李建生2, 杨小红2, 白光红1,*()
), 胡书婷2(), 张凯2, 崔则瑾2, 李建生2, 杨小红2, 白光红1,*()
Xin-Ran SONG1(), Shu-Ting HU2(), Kai ZHANG2, Ze-Jin CUI2, Jian-Sheng LI2, Xiao-Hong YANG2, Guang-Hong BAI1,*()
摘要:
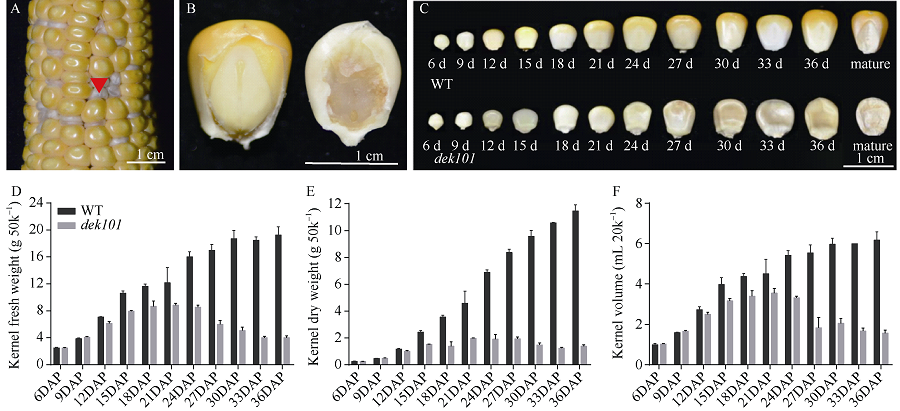
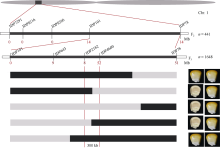
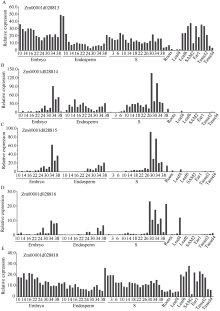
籽粒作为玉米储藏器官, 其发育程度和物质储存直接影响玉米的产量和品质。本研究在玉米双单倍体系选育过程中发现可稳定遗传的籽粒缺陷突变体, 命名为defective kernel 101 (dek101)。该突变体籽粒皱缩, 粒重显著降低, 胚致死, 胚乳发育缺陷, 不能成苗。在授粉后12 d, dek101开始出现明显的发育异常, 授粉后21 d籽粒鲜重、干重、体积不再增加。扫描电镜观察发现, 与野生型相比, dek101淀粉粒显著变小。遗传分析证实该突变性状受隐性单基因控制。利用441个F2单株和1648个F3单株, 将该基因定位在1号染色体的标记IDP2182和IDP4600之间, 物理区间约300 kb, 共有5个预测基因。这些结果为挖掘与玉米籽粒发育有关的功能基因, 解析籽粒发育机制奠定了基础。
| [1] | Rossi V, Hartings H, Thompson R, Mario M . Genetic and molecular approaches for upgrading starch and protein fractions in maize kernels. Maydica, 2001,46:147-158. |
| [2] |
Shannon J C, Pien F M, Cao H, Liu K C . Brittle-1, an adenylate translocator, facilitates transfer of extraplastidial synthesized ADP-Glucose into amyloplasts of maize endosperms. Plant Physiol, 1998,117:1235-1252.
doi: 10.1104/pp.117.4.1235 pmid: 9701580 |
| [3] |
Dickinson D B, Preiss J . Presence of ADP-Glucose pyrophosphorylase in shrunken-2 and brittle-2 mutants of maize endosperm. Plant Physiol, 1969,44:1058-1062.
doi: 10.1104/pp.44.7.1058 pmid: 16657157 |
| [4] |
Zhang X L, Colleoni C, Ratushna V, Colleoni M S, Martha J, Myers A . Molecular characterization demonstrates that the Zea mays gene sugary2 codes for the starch synthase isoform SSIIa. Plant Mol Biol, 2004,54:865-879.
doi: 10.1007/s11103-004-0312-1 |
| [5] | Wang Y J, White P, Pollak L, Jane J L . Characterization of starch structure of 17 maize endosperm mutant genotypes with Oh43 inbred line background. Cereal Chem, 1993,70:171-179. |
| [6] |
Shure M, Wessler S, Fedoroff N . Molecular identification and isolation of the waxy locus in maize. Cell, 1983,35:225-233.
doi: 10.1016/0092-8674(83)90225-8 pmid: 6313224 |
| [7] |
Fu S N, Meeley R, Scanlon M J . Empty pericarp2 encodes a negative regulator of the heat shock response and is required for maize embryogenesis. Plant Cell, 2002,14:3119-3132.
doi: 10.1105/tpc.006726 pmid: 12468731 |
| [8] |
Jose F, Gutierrez M, Mauro D P . Empty pericarp4 encodes a mitochondrion-targeted Pentatricopeptide repeat protein necessary for seed development and plant growth in maize. Plant Cell, 2007,19:196-210.
doi: 10.1105/tpc.105.039594 pmid: 17259266 |
| [9] |
He Y H, Wang J G, Qi W W, Song R T . Maize dek15 encodes the cohesin-loading complex subunit SCC4 and is essential for chromosome segregation and kernel development. Plant Cell, 2019,31:465-485.
doi: 10.1105/tpc.18.00921 pmid: 30705131 |
| [10] |
Garcia N, Li Y B, Dooner H K, Messing J . Maize defective kernel mutant generated by insertion of a Ds element in a gene encoding a highly conserved TTI2 cochaperone. Proc Natl Acad Sci USA, 2017,114:5165-5170.
doi: 10.1073/pnas.1703498114 pmid: 28461460 |
| [11] |
Qi W W, Lu L, Huang S C, Song R T . Maize dek44 encodes mitochondrial ribosomal protein L9 and is required for seed development. Plant Physiol, 2019,180:2106-2119.
doi: 10.1104/pp.19.00546 pmid: 31182559 |
| [12] |
Shen Y, Li C L, McCarty D R, Meeley R, Bao C T . Embryo defective 12 encodes the plastid initiation factor 3 and is essential for embryogenesis in maize. Plant J, 2013,74:792-804.
doi: 10.1111/tpj.12161 |
| [13] |
Li C, Shen Y, Meeley R, McCarty D R, Bao C T . Embryo defective 14 encodes a plastid-targeted cGTPase essential for embryogenesis in maize. Plant J, 2015,84:785-799.
doi: 10.1111/tpj.13045 pmid: 26771182 |
| [14] |
Mertz ET, Bates L S, Nelson O E . Mutant gene that changes protein composition and increases lysine content of maize endosperm. Science, 1964,145:279-280.
doi: 10.1126/science.145.3629.279 pmid: 14171571 |
| [15] |
Ueda T, Waverczak W, Ward K, Sher K, Ketuda M, Schmidt R J, Messing J . Mutations of the 22- and 27-kD zein promoters affect transactivation by the Opaque-2 protein. Plant Cell, 1992,4:701-709.
doi: 10.1105/tpc.4.6.701 pmid: 1392591 |
| [16] | Coleman C E, Larkins B A . The Prolamins of Maize. Seed Proteins. Springer Netherlands, 1999. pp 109-139. |
| [17] | Nelson O E, Mertz E T, Bates L S . Second mutant gene affecting the amino acid pattern of maize. Endosperm Proteins, 1966,150:1469-1470. |
| [18] |
Huang Y C, Wang H H, Huang X, Wang Q, Wang C J, An D, Li J Q, Wang W Q, Wu Y R . Maize VKS1 regulates mitosis and cytokinesis during early endosperm development. Plant Cell, 2019,31:1238-1256.
doi: 10.1105/tpc.18.00966 pmid: 30962394 |
| [19] |
Zheng X X, Li Q, Li C S, An D, Xiao Q, Wang W Q, Wu Y R . Intra-kernel reallocation of proteins in maize depends on VP1-mediated scutellum development and nutrient assimilation. Plant Cell, 2019,31:2613-2635.
doi: 10.1105/tpc.19.00444 pmid: 31530735 |
| [20] |
Chen J, Zeng B, Zhang M, Xie S J, Wang G K, Hauck A, Lai J S . Dynamic transcriptome landscape of maize embryo and endosperm development. Plant Physiol, 2014,166:252-264.
doi: 10.1104/pp.114.240689 |
| [21] |
Chourey P S, Nelson O E . The enzymatic deficiency conditioned by the shrunken-1, mutations in maize. Biochem Genet, 1976,14:1041-1055.
doi: 10.1007/BF00485135 pmid: 1016220 |
| [22] |
Hannah L C, Jr O E N . Characterization of ADP-glucose pyrophosphorylase from shrunken-2, and brittle-2, mutants of maize. Biochemical Genet, 1976,14:547-560.
doi: 10.1007/BF00485834 |
| [23] |
Gao M, Wanat J, Stinard P S, James M G, Myers A M . Characterization of dull1, a maize gene coding for a novel starch synthase. Plant Cell, 1998,10:399-412.
doi: 10.1105/tpc.10.3.399 pmid: 9501113 |
| [24] |
Magnard J L, Heckel T, Massonneau A, Wisniewski J P, Cordelier S, Lassagne H, Perez P, Dumas C, Rogowsky P M . Morphogenesis of maize embryos requires ZmPRPL35-1 encoding a plastid ribosomal protein. Plant Physiol, 2004,134:649-663.
doi: 10.1104/pp.103.030767 pmid: 14730079 |
| [25] |
Qi W, Tian Z, Lu L, Chen X, Chen X, Zhang W, Song R T . Editing of mitochondrial transcripts nad3 and cox2 by dek10 is essential for mitochondrial function and maize plant development. Genetics, 2017,205:1489-1501.
doi: 10.1534/genetics.116.199331 pmid: 28213476 |
| [26] |
Chen X, Feng F, Qi W, Xu L, Yao D, Wang Q, Song R T . Dek35 encodes a PPR protein that affects cis-splicing of mitochondrial nad4 intron 1 and seed development in maize. Mol Plant, 2017,10:427-441.
doi: 10.1016/j.molp.2016.08.008 pmid: 27596292 |
| [27] |
Wang G, Zhong M, Shuai B, Song J, Zhang J, Han L, Ling H, Tang Y, Wang G, Song R T . E+ subgroup PPR protein defective kernel 36 is required for multiple mitochondrial transcripts editing and seed development in maize and Arabidopsis. New Phytol, 2017,214:1563-1578.
doi: 10.1111/nph.14507 pmid: 28277611 |
| [28] |
Dai D, Luan S, Chen X, Wang Q, Feng Y, Zhu C, Qi W, Song R T . Maize dek37 encodes a P-type PPR protein that affects cis-splicing of mitochondrial nad2 intron 1 and seed development. Genetics, 2018,208:1069-1082.
doi: 10.1534/genetics.117.300602 pmid: 29301905 |
| [29] |
Li X, Gu W, Sun S, Chen Z, Chen J, Song W, Zhao H, Lai J . Defective kernel 39 encodes a PPR protein required for seed development in maize. J Integr Plant Biol, 2018,60:45-64.
doi: 10.1111/jipb.12602 |
| [30] |
Ren R C, Lu X, Zhao Y J, Wei Y M, Wang L L, Zhang L, Zhang W T, Zhang C, Zhang X S, Zhao X Y . Pentatricopeptide repeat protein DEK40 is required for mitochondrial function and kernel development in maize. J Exp Bot, 2019,70:6163-6179.
doi: 10.1093/jxb/erz391 pmid: 31598687 |
| [31] |
Zuo Y, Feng F, Qi W W, Song R T . Dek42 encodes an RNA-binding protein that affects alternative pre-mRNA splicing and maize kernel development. J Integr Plant Biol, 2019,61:728-748.
doi: 10.1111/jipb.12798 |
| [32] |
Stein E L, Darren G, Rudolf J, Jennifer A L, Evgueni A, Mark C, Niu X M, Meeley R, Nichols S, Olsen O A . The defective kernel 1 (dek 1) gene required for aleurone cell development in the endosperm of maize grains encodes a membrane protein of the calpain gene superfamily. Proc Natl Acad Sci USA, 2002,99:5460-5465.
doi: 10.1073/pnas.042098799 pmid: 11929961 |
| [33] |
Zhu C, Jin G, Fang P, Zhang Y, Feng X, Tang Y, Qi W W, Song R T . Maize Pentatricopeptide repeat protein DEK41 affects cis-splicing of mitochondrial nad4 intron 3 and is required for normal seed development. J Exp Bot, 2019,70:3795-3808.
doi: 10.1093/jxb/erz193 pmid: 31020318 |
| [34] |
Yi G, Lauter A M, Scott P, Becraft P W . The thick aleurone1 mutant defines a negative regulation of maize aleurone cell fate that functions downstream of defective kernel 1. Plant Physiol, 2011,156:1826-1836.
doi: 10.1104/pp.111.177725 |
| [35] |
Xu C, Min J R . Structure and function of WD40 domain proteins. Protein Cell, 2011,2:202-214.
doi: 10.1007/s13238-011-1018-1 |
| [36] |
Xie Y R, Chen Z Y, Brown R L, Bhatnagar D . Expression and functional characterization of two pathogenesis-related protein 10 genes from Zea mays. J Plant Physiol, 2010,167:121-130.
doi: 10.1016/j.jplph.2009.07.004 pmid: 19682768 |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [4] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [5] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [6] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [7] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [8] | 王好让, 张勇, 于春淼, 董全中, 李微微, 胡凯凤, 张明明, 薛红, 杨梦平, 宋继玲, 王磊, 杨兴勇, 邱丽娟. 大豆突变体ygl2黄绿叶基因的精细定位[J]. 作物学报, 2022, 48(4): 791-800. |
| [9] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [10] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [11] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [12] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [13] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [14] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [15] | 张倩, 韩本高, 张博, 盛开, 李岚涛, 王宜伦. 控失尿素减施及不同配比对夏玉米产量及氮肥效率的影响[J]. 作物学报, 2022, 48(1): 180-192. |
|
||