欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (12): 1839-1849.doi: 10.3724/SP.J.1006.2020.04075
张之昊1,2( ), 王俊1, 刘章雄2,*(), 邱丽娟1,2,*()
), 王俊1, 刘章雄2,*(), 邱丽娟1,2,*()
ZHANG Zhi-Hao1,2(), WANG Jun1, LIU Zhang-Xiong2,*(), QIU Li-Juan1,2,*()
摘要:
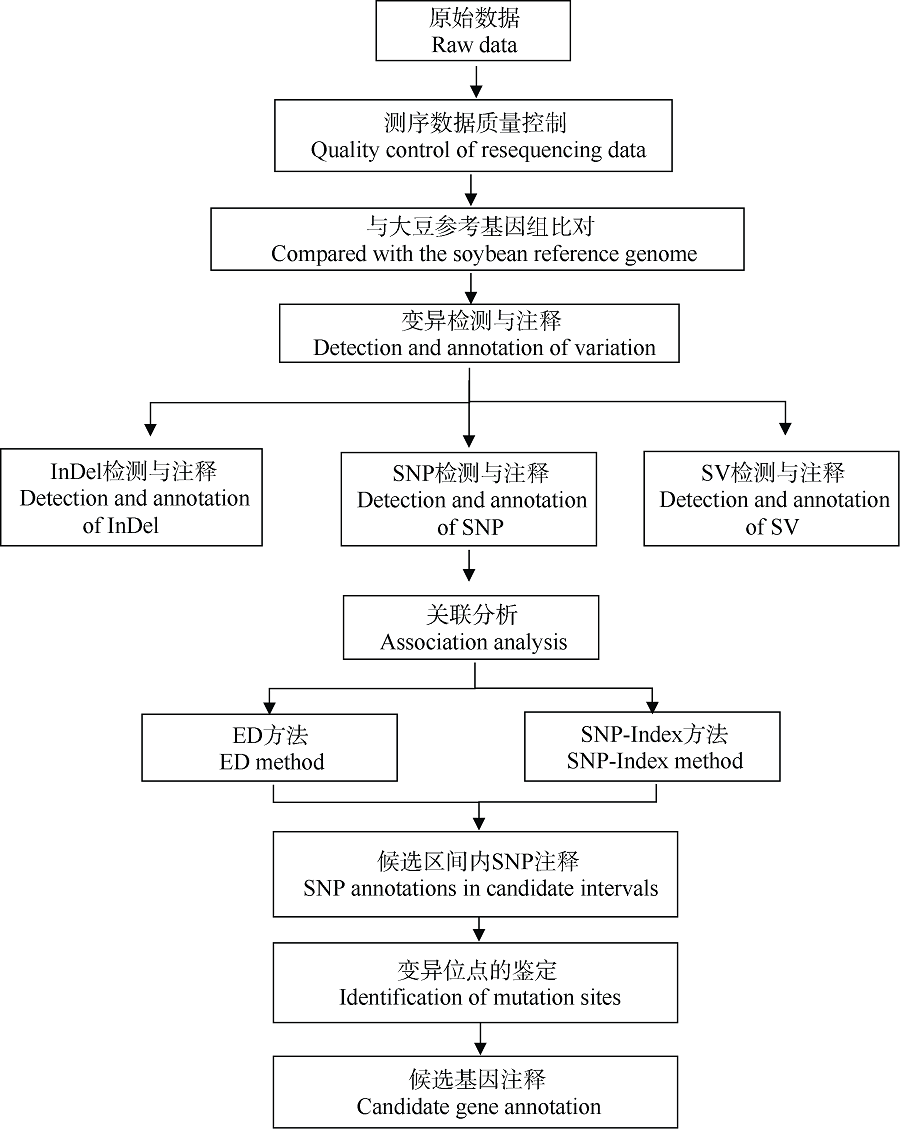
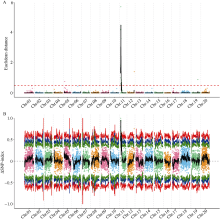
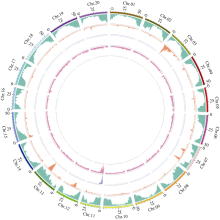
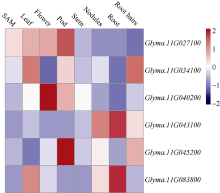
栽培大豆(Glycine max)叶片一般为三出复叶, 也有个别品种或植株突变体产生4~7个小叶, 为多小叶。复叶的形成使植物对外界环境的适应能力增强, 对大豆多小叶相关基因的挖掘和研究有助于改善大豆农艺性状和产量表现。本研究从大豆栽培品种中品661的突变体库中鉴定出一个多小叶突变体——中黄622, 每个复叶有4~9个小叶。利用该突变体与中品661配制组合, 分别于北京和海南调查F2和F2:3植株叶片表型, 结果表明, 多小叶性状受1对不完全显性基因控制。采用BSA-seq方法进行定位, 利用F2正常三出复叶和多小叶个体分别构建混池, 测序结果与参考基因组平均比对效率为98.83%, 平均覆盖深度为32.75×, 基因组覆盖度为99.22%。ED方法关联分析发现, 在11号染色体定位到2个区域, 总长度为5.29 Mb, 共包含1103个基因。根据SNP-index方法关联分析, 当置信度为0.99时, 在11号染色体鉴定出3个区域, 总长度为3.42 Mb, 共包含701个基因。2种关联分析方法同时定位的基因有690个, 亲本之间存在SNP的基因有6个。本研究结果为大豆多小叶基因图位克隆奠定了基础。
| [1] |
Vogel S . Leaves in the lowest and highest winds: temperature, force and shape. New Phytol, 2009,183:13-26.
doi: 10.1111/j.1469-8137.2009.02854.x pmid: 19413689 |
| [2] | 宗春美, 岳岩磊, 邵广忠, 童淑媛, 徐显利, 杜震宇, 任海祥 . 多小叶源对大豆光合特性和产量的影响. 大豆科学, 2010,29:627-626. |
| Zong C M, Yue Y L, Shao G Z, Tong S Y, Xu X L, Du Z Y, Ren H X . Effects of multifoliolate compound leaf on photosynthetic characteristics and yield of soybean. Soybean Sci, 2010,29:627-626 (in Chinese with English abstract). | |
| [3] |
Fehr W R . Genetic control of leaflet number in soybeans. Crop Sci, 1972,12:221-224.
doi: 10.2135/cropsci1972.0011183X001200020023x |
| [4] |
Jeong S C, Kim J H, Bae D N . Genetic analysis of the Lf1 gene that controls leaflet number in soybean. Theor Appl Genet, 2017,130:1685-1692.
doi: 10.1007/s00122-017-2918-0 pmid: 28516383 |
| [5] |
Devine T E . The Pd2 and Lf2 loci define soybean linkage group 16. Crop Sci, 2003,43:2028-2030.
doi: 10.2135/cropsci2003.2028 |
| [6] |
Seversike T M, Ray J D, Shultz J L, Purcell L C . Soybean molecular linkage group B1 corresponds to classical linkage group 16 based on map location of the lf2 gene. Theor Appl Genet, 2008,117:143-147.
doi: 10.1007/s00122-008-0759-6 |
| [7] | 傅来卿 . 大豆双复叶和多小叶突变体的研究. 大豆科学, 1986,5:283-288. |
| Fu L Q . Study of mutants with opposite trifoliate leaves and multi-leaflet leaves in soybean. Soybean Sci, 1986,5:283-288 (in Chinese with English abstract). | |
| [8] | 王克晶, 李福山, 周涛, 许占有 . 来源于野生大豆的多小叶性状遗传分析. 大豆科学, 2001,20:22-25. |
| Wang K J, Li F S, Zhou T, Xu Z Y . Inheritance of a five leaflet character arising from wild soybean (Glycine soja Sieb. et Zucc.) in soybeans (G. max (L.) Merr.). Soybean Sci, 2001,20:22-25 (in English with Chinse abstract). | |
| [9] |
Nawy T, Bayer M, Mravec J, Friml J, Birnbaum K D, Lukowitz W . The GATA factor HANABA TARANU is required to position the proembryo boundary in the early Arabidopsis embryo. Dev Cell, 2010,19:103-113.
doi: 10.1016/j.devcel.2010.06.004 pmid: 20643354 |
| [10] | Gallavotti A, Long J A, Stanfield S, Yang X, Jackson D, Vollbrecht E, Schmidt R J . The control of axillary meristem fate in the maize ramosa pathway. Development(Cambridge, England), 2010,137:2849-2856. |
| [11] |
Vlad D, Kierzkowski D, Rast M I, Vuolo F, Ioio R D, Galinha C, Gan X, Hajheidari M, Hay A, Smith R S, Huijser P, Bailey C D, Tsiantis M . Leaf shape evolution through duplication, regulatory diversification, and loss of a homeobox gene. Science, 2014,343:780-783.
doi: 10.1126/science.1248384 pmid: 24531971 |
| [12] |
Stewart G C, Roeder A H K, Patrick S, Chris S, Wolfgang L, Hector C . A genetic screen for mutations affecting cell division in the Arabidopsis thaliana embryo identifies seven loci required for cytokinesis. PLoS One, 2016,11:e0146492.
doi: 10.1371/journal.pone.0146492 pmid: 26745275 |
| [13] |
Abe A, Kosugi S, Yoshida K, Natsume S, Takagi H, Kanzaki H, Matsumura H, Yoshida K, Mitsuoka C, Tamiru M, Innan H, Cano L, Kamoun S, Terauchi R . Genome sequencing reveals agronomically important loci in rice using MutMap. Nat Biotechnol, 2012,30:174-178.
doi: 10.1038/nbt.2095 pmid: 22267009 |
| [14] |
Abe A1, Kosugi S, Yoshida K, Natsume S, Takagi H, Kanzaki H, Matsumura H, Yoshida K, Mitsuoka C, Tamiru M, Innan H, Cano L, Kamoun S, Terauchi R . QTL-seq: rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J, 2013,74:174-183.
doi: 10.1111/tpj.12105 |
| [15] |
Zhang H, Wang X, Pan Q, Li P, Liu Y, Lu X, Zhong W, Li M, Han L, Li J, Wang P, Li D, Liu Y, Li Q, Yang F, Zhang Y M, Wang G, Li L . QTG-Seq accelerates QTL fine mapping through QTL partitioning and whole-genome sequencing of bulked segregant samples. Mol Plant, 2019,12:426-437.
doi: 10.1016/j.molp.2018.12.018 pmid: 30597214 |
| [16] | Klein H, Xiao Y, Conklin P A, Govindarajulu R, Kelly J A, Scanlon M J, Whipple C J, Bartlett M . Bulked-segregant analysis coupled to whole genome sequencing (BSA-Seq) for rapid gene cloning in maize. G3: Genes Genomes Genet, 2018,8:3583-3592. |
| [17] |
Song Q J, Jenkins J, Jia G F, Hyten D L, Pantalone V, Jackson S A . Construction of high resolution genetic linkage maps to improve the soybean genome sequence assembly Glyma1.01. BMC Genomics, 2016,17:33.
doi: 10.1186/s12864-015-2344-0 pmid: 26739042 |
| [18] |
McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K, Kernytsky A, Garimella K, Altshuler D, Gabriel S, Daly M, DePristo M A , The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res, 2010,20:1297-1303.
doi: 10.1101/gr.107524.110 pmid: 20644199 |
| [19] |
Cingolani P, Platts A, Wang L L, Coon M, Nguyen T, Wang L, Land S J, Lu X Y, Ruden D M . A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly, 2012,6:80-92.
doi: 10.4161/fly.19695 |
| [20] |
Hill J T, Demarest B L, Bisgrove B W, Gorsi B, Su Y C, Yost H J . MMAPPR: mutation mapping analysis pipeline for pooled RNA-seq. Genome Res, 2013,23:687-697.
doi: 10.1101/gr.146936.112 |
| [21] |
Fekih R, Takagi H, Tamiru M, Abe A, Natsume S, Yaegashi H, Sharma S, Sharma S, Kanzaki H, Matsumura H, Saitoh H, Mitsuoka C, Utsushi H, Uemura A, Kanzaki E, Kosugi S, Yoshida K, Cano L, Kamoun S, Terauchi R . MutMap+: Genetic mapping and mutant identification without crossing in rice. PLoS One, 2013,8:e68529.
doi: 10.1371/journal.pone.0068529 pmid: 23874658 |
| [22] |
Altschul S F, Madden T L, Schäffer A A, Zhang J, Zhang Z, Miller W, Lipman D J . Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res, 1997,25:3389-3402.
doi: 10.1093/nar/25.17.3389 pmid: 9254694 |
| [23] | Deng Y, Li J Q, Wu S F, Zhu Y P, Chen Y W, He F C . Integrated nr database in protein annotation system and its localization. Comput Engineer, 2006,32:71-72. |
| [24] |
Ashburner M, Ball C A, Blake J A, Botstein D, Butler H, Cherry M, Davis A P, Dolinski K, Dwight S S, Eppig J T, Harris M A, Hill D P, Issel-Tarver L, Kasarskis A, Lewis S, Matese J C, Richardson J E, Ringwald M, Rubin G M, Sherlock G . Gene ontology: tool for the unification of biology. Nat Genet, 2000,25:25-29.
doi: 10.1038/75556 pmid: 10802651 |
| [25] |
Kanehisa M, Goto S, Kawashima S, Okuno Y, Hattori M . The KEGG resource for deciphering the genome. Nucleic Acids Res, 2004,32:D277-D280.
doi: 10.1093/nar/gkh063 pmid: 14681412 |
| [26] |
Tatusov R L, Galperin M Y, Natale D A, Koonin E V . The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res, 2000,28:33-36.
doi: 10.1093/nar/28.1.33 pmid: 10592175 |
| [27] | 杨霞, 高金珊, 杨素欣 . 豆科复叶发育分子遗传机制的研究进展. 植物生理学报, 2017,53:905-915. |
| Yang X, Gao J S, Yang S X . Progress of molecular mechanism of compound leaf development in legume plants. Plant Physiol Commun, 2017,53:905-915 (in Chinese with English abstract). | |
| [28] |
Long J A, Moan E I, Medford J I, Barton M K . A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature (London), 1996,379:66-69.
doi: 10.1038/379066a0 |
| [29] |
Bharathan G, Goliber T E, Moore C, Kessler S, Pham T, Sinha N R . Homologies in leaf form inferr ed from KNOXI gene expression during development. Science, 2002,296:1858-1860.
doi: 10.1126/science.1070343 pmid: 12052958 |
| [30] |
Lincoln C, Long J, Yamaguchi J, Serikawa K, Hake S . A knotted1-like homeobox gene in Arabidopsis is expressed in the vegetative meristem and dramatically alters leaf morphology when overexpressed in transgenic plants. Plant Cell, 1994,6:1859-1876.
doi: 10.1105/tpc.6.12.1859 pmid: 7866029 |
| [31] |
Hay A, Tsiantis M . The genetic basis for differences in leaf form between Arabidopsis thaliana and its wild relative Cardamine hirsuta. Nat Genet, 2006,38:942-947.
doi: 10.1038/ng1835 pmid: 16823378 |
| [32] |
Shani E, Burko Y, Ben-Yaakov L, Berger Y, Amsellem Z, Goldshmidt A, Sharon E, Ori N . Stage-specific regulation of Solanum lycopersicum leaf maturation by class 1 KNOTTED1- LIKE HOMEOBOX proteins. Plant Cell, 2009,21:3078-3092.
doi: 10.1105/tpc.109.068148 pmid: 19820191 |
| [33] |
Hareven D, Gutfinger T, Parnis A, Eshed Y, Lifschitz E . The making of a compound leaf: genetic manipulation of leaf architecture in tomato. Cell, 1996,84:735-744.
doi: 10.1016/s0092-8674(00)81051-x pmid: 8625411 |
| [34] |
Byrne M E, Barley R, Curtis M, Arroyo J M, Dunham M, Hudson A, Martienssen R A . Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Nature, 2000,408:967-971.
doi: 10.1038/35050091 pmid: 11140682 |
| [35] |
Waites R, Selvadurai H R N, Oliver I R, Hudson A . The PHANTASTICA gene encodes a MYB transcription factor involved in growth and dorsoventrality of lateral organs in Antirrhinum. Cell, 1998,93:779-789.
doi: 10.1016/s0092-8674(00)81439-7 pmid: 9630222 |
| [36] |
Kim M, Pham T, Hamidi A, McCormick S, Kuzoff R K, Sinha N . Reduced leaf complexity in tomato wiry mutants suggests a role for PHAN and KNOX genes in generating compound leaves. Development, 2003,130:4405-4415.
doi: 10.1242/dev.00655 pmid: 12900456 |
| [37] |
Taylor S, Hofer J, Murfet I . Stamina pistilloida, the pea ortholog of Fim and UFO, is required for normal development of flowers, inflorescences, and leaves. Plant Cell, 2001,13:31-46.
doi: 10.1105/tpc.13.1.31 pmid: 11158527 |
| [38] |
Dong Z C, Zhao Z, Liu C W, Luo J H, Yang J, Huang W H, Hu X H, Wang T L, Luo D . Floral patterning in Lotus japonicus. Plant Physiol, 2005,137:1272-1282.
doi: 10.1104/pp.104.054288 pmid: 15824286 |
| [39] |
Jiang F K, Guo M, Yang F, Duncan K, Jackson D, Rafalski A, Wang S C, Li B L . Mutations in an AP2 transcription factor- like gene affect internode length and leaf shape in maize. PLoS One, 2012,7:e37040.
doi: 10.1371/journal.pone.0037040 pmid: 22649507 |
| [40] |
Soll M D, Ibba T M . Aminoacyl-tRNA synthesis. Annu Rev Biochem, 2000,69:617-650.
doi: 10.1146/annurev.biochem.69.1.617 pmid: 10966471 |
| [41] |
Fatscher T, Boehm V, Gehring N H . Mechanism, factors, and physiological role of nonsense-mediated mRNA decay. Cell Mol Life Sci, 2015,72:4523-4544.
doi: 10.1007/s00018-015-2017-9 pmid: 26283621 |
| [42] | 柴宝峰, 王美, 石文鑫, 柴杨丽, 吕佳 . 无义mRNA降解途径的机制与进化. 山西大学学报(自然科学版), 2017,40:639-644. |
| Chai B F, Wang M, Shi W X, Chai Y L, Lyu J . Mechanism and evolution of nonsense-mediated mRNA decay. J Shanxi Univ (Nat Sci Edn), 2017,40:639-644 (in Chinese with English abstract). | |
| [43] | 贾晓波, 胡剑 . 无义介导的mRNA降解. 中国生物化学与分子生物学报, 2012,28(2):22-27. |
| Jia X B, Hu J . Nonsense-mediated mRNA decay. Chin J Biochem Mol Biol, 2012,28(2):22-27 (in Chinese with English abstract). | |
| [44] |
Yamashita A . Role of SMG-1-mediated Upf1 phosphorylation in mammalian nonsense-mediated mRNA decay. Genes Cells, 2013,18:161-175.
doi: 10.1111/gtc.12033 |
| [45] |
Bhattacharya A, Köhrer C, Mandal D, Rajbhandary U L . Nonsense suppression in archaea. Proc Natl Acad Sci USA, 2015,112:6015-6020.
doi: 10.1073/pnas.1501558112 pmid: 25918386 |
| [46] | 肖朝文, 陈福禄, 傅永福 . AT-hook基因AHL27过量表达延迟拟南芥开花. 中国农业科技导报, 2009,11(4):93-98. |
| Xiao C W, Chen F L, Fu Y F . Over-expression of AT-hook gene AHL27 can delay the flowering of Arabidopsis. J Agric Sci Technol, 2009,11(4):93-98 (in Chinese with English abstract). | |
| [47] |
Getzenberg R H, Pienta K J, Ward W S, Coffey D S . Nuclear structure and the three-dimensional organization of DNA. J Cell Biochem, 1991,47:289-299.
doi: 10.1002/jcb.240470402 pmid: 1795013 |
| [1] | 陈玲玲, 李战, 刘亭萱, 谷勇哲, 宋健, 王俊, 邱丽娟. 基于783份大豆种质资源的叶柄夹角全基因组关联分析[J]. 作物学报, 2022, 48(6): 1333-1345. |
| [2] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [3] | 王炫栋, 杨孙玉悦, 高润杰, 余俊杰, 郑丹沛, 倪峰, 蒋冬花. 拮抗大豆斑疹病菌放线菌菌株的筛选和促生作用及防效研究[J]. 作物学报, 2022, 48(6): 1546-1557. |
| [4] | 于春淼, 张勇, 王好让, 杨兴勇, 董全中, 薛红, 张明明, 李微微, 王磊, 胡凯凤, 谷勇哲, 邱丽娟. 栽培大豆×半野生大豆高密度遗传图谱构建及株高QTL定位[J]. 作物学报, 2022, 48(5): 1091-1102. |
| [5] | 李阿立, 冯雅楠, 李萍, 张东升, 宗毓铮, 林文, 郝兴宇. 大豆叶片响应CO2浓度升高、干旱及其交互作用的转录组分析[J]. 作物学报, 2022, 48(5): 1103-1118. |
| [6] | 彭西红, 陈平, 杜青, 杨雪丽, 任俊波, 郑本川, 罗凯, 谢琛, 雷鹿, 雍太文, 杨文钰. 减量施氮对带状套作大豆土壤通气环境及结瘤固氮的影响[J]. 作物学报, 2022, 48(5): 1199-1209. |
| [7] | 王好让, 张勇, 于春淼, 董全中, 李微微, 胡凯凤, 张明明, 薛红, 杨梦平, 宋继玲, 王磊, 杨兴勇, 邱丽娟. 大豆突变体ygl2黄绿叶基因的精细定位[J]. 作物学报, 2022, 48(4): 791-800. |
| [8] | 杜晓芬, 王智兰, 韩康妮, 连世超, 李禹欣, 张林义, 王军. 谷子叶绿体基因RNA编辑位点的鉴定与分析[J]. 作物学报, 2022, 48(4): 873-885. |
| [9] | 李瑞东, 尹阳阳, 宋雯雯, 武婷婷, 孙石, 韩天富, 徐彩龙, 吴存祥, 胡水秀. 增密对不同分枝类型大豆品种同化物积累和产量的影响[J]. 作物学报, 2022, 48(4): 942-951. |
| [10] | 杜浩, 程玉汉, 李泰, 侯智红, 黎永力, 南海洋, 董利东, 刘宝辉, 程群. 利用Ln位点进行分子设计提高大豆单荚粒数[J]. 作物学报, 2022, 48(3): 565-571. |
| [11] | 周悦, 赵志华, 张宏宁, 孔佑宾. 大豆紫色酸性磷酸酶基因GmPAP14启动子克隆与功能分析[J]. 作物学报, 2022, 48(3): 590-596. |
| [12] | 王娟, 张彦威, 焦铸锦, 刘盼盼, 常玮. 利用PyBSASeq算法挖掘大豆百粒重相关位点与候选基因[J]. 作物学报, 2022, 48(3): 635-643. |
| [13] | 董衍坤, 黄定全, 高震, 陈栩. 大豆PIN-Like (PILS)基因家族的鉴定、表达分析及在根瘤共生固氮过程中的功能[J]. 作物学报, 2022, 48(2): 353-366. |
| [14] | 张国伟, 李凯, 李思嘉, 王晓婧, 杨长琴, 刘瑞显. 减库对大豆叶片碳代谢的影响[J]. 作物学报, 2022, 48(2): 529-537. |
| [15] | 赵改会, 李书宇, 詹杰鹏, 李晏斌, 师家勤, 王新发, 王汉中. 甘蓝型油菜角果数突变体基因的定位及候选基因分析[J]. 作物学报, 2022, 48(1): 27-39. |
|
||