欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (6): 1389-1400.doi: 10.3724/SP.J.1006.2022.12035
郑崇珂1( ), 周冠华1, 牛淑琳1,2, 和亚男1, 孙伟1, 谢先芝1,*()
), 周冠华1, 牛淑琳1,2, 和亚男1, 孙伟1, 谢先芝1,*()
ZHENG Chong-Ke1(), ZHOU Guan-Hua1, NIU Shu-Lin1,2, HE Ya-Nan1, SUN wei1, XIE Xian-Zhi1,*()
摘要:
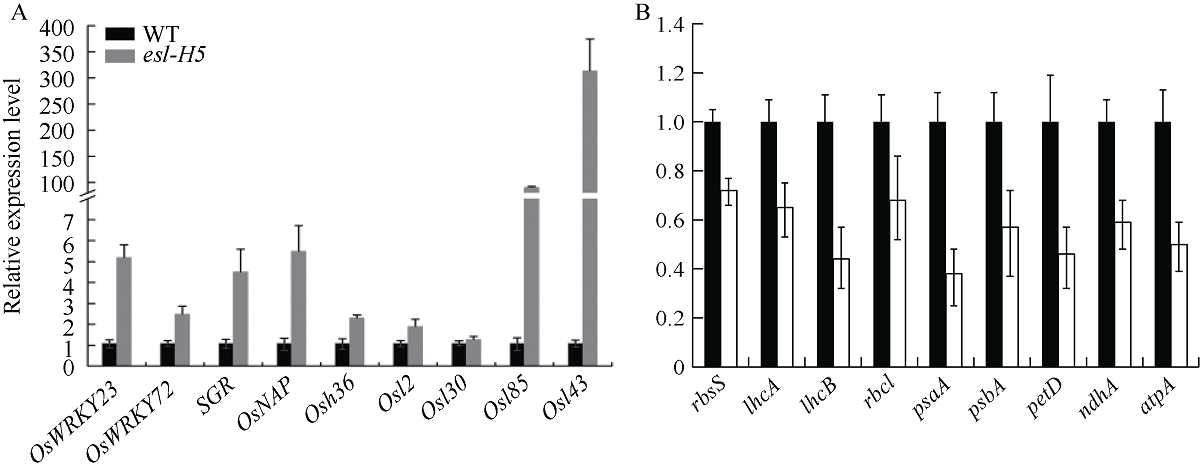
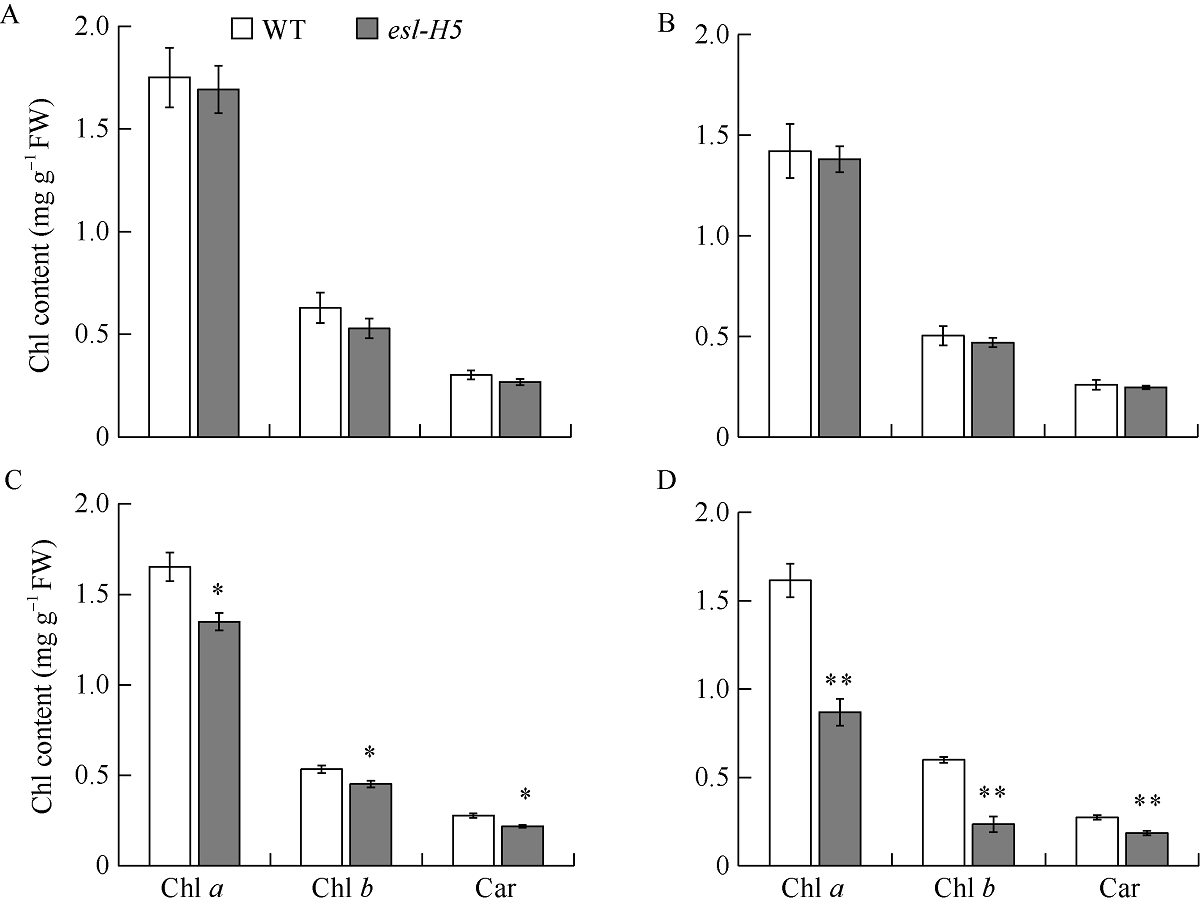
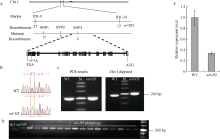
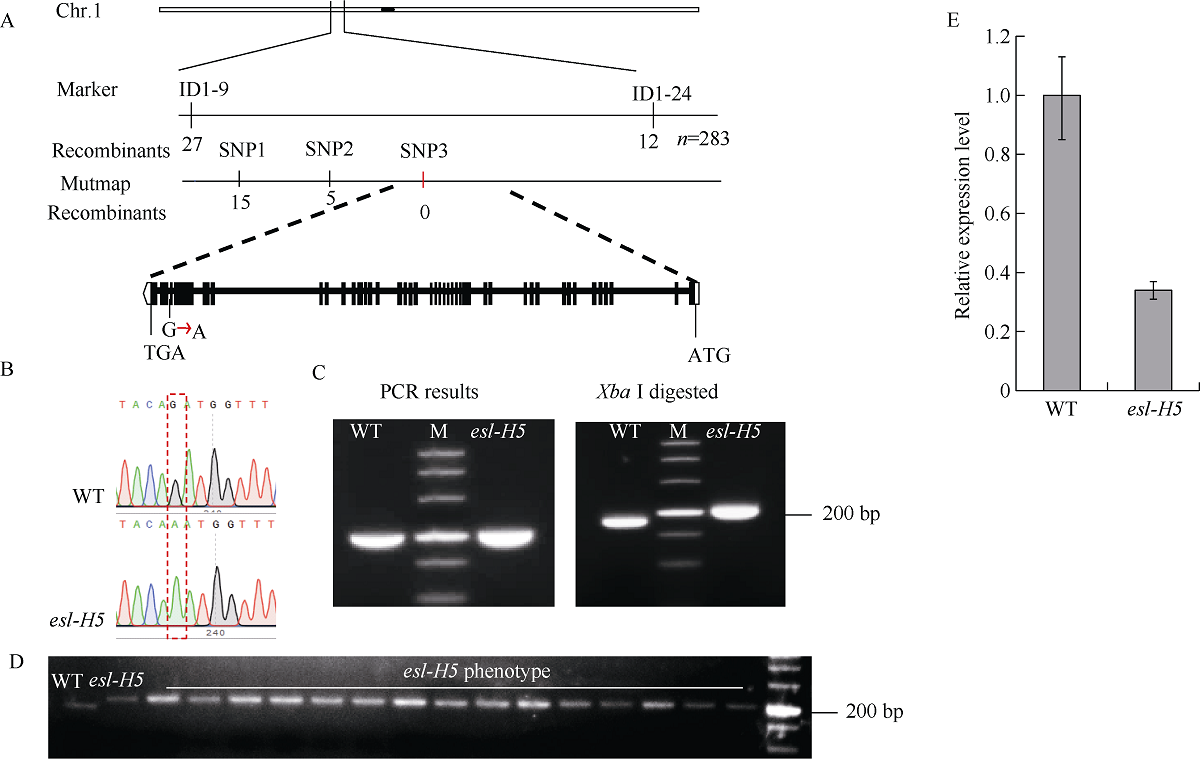
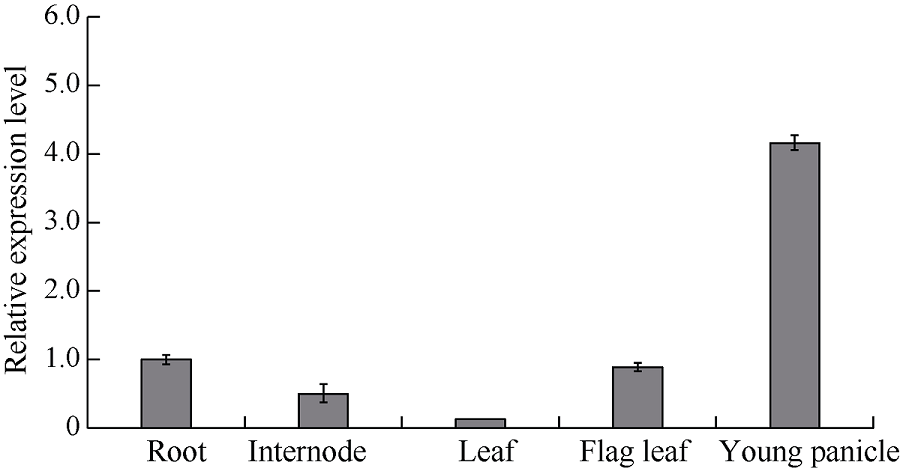
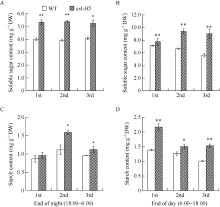
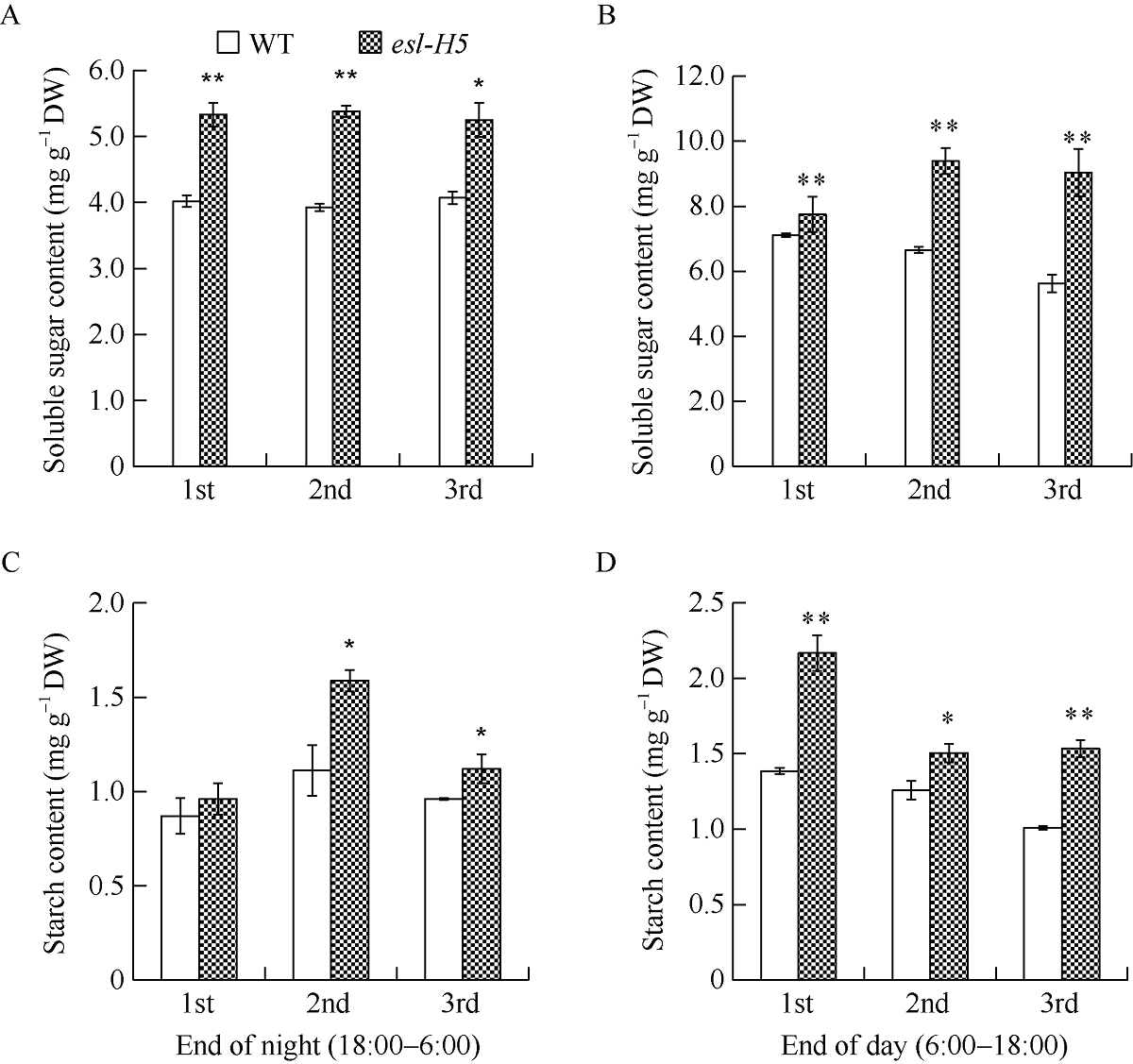
在甲基磺酸乙酯(EMS)诱变粳稻淮稻5号的突变体库中, 筛选到一个稳定遗传的叶片黄化早衰突变体esl-H5 (early senescence leaf H5)。该突变体幼苗期表型正常, 播种后50 d开始出现下部叶片黄化早衰表型。与野生型相比, esl-H5突变体抽穗期延迟, 株高、穗长、穗粒数、有效分蘖数、千粒重等均显著降低, 叶绿素含量显著减少。遗传分析表明该突变受一对隐性基因调控。分子标记定位结果显示, 该突变基因定位于1号染色体。通过MutMap分析发现编码胼胝质合成酶基因Os01g0533000的最后一个外显子内有一个碱基G变成了A, 这导致翻译提前终止。进化树分析结果显示, ESL-H5与拟南芥AtGSL7 (Glucan Synthase-Like 7)同源性最高。糖含量测定表明esl-H5突变体中可溶性糖和淀粉含量增高, 推测ESL-H5功能缺失后影响了光合产物的转运, 导致叶片中糖含量显著增高, 进而引起叶片衰老。qRT-PCR结果显示, esl-H5突变体中抗病相关基因PR1a、PR1b、PR2、PR4、PR5和PR10表达量均高于野生型, 这与突变体的白叶枯病抗性明显提高一致。上述研究结果为研究糖信号调控水稻衰老和抗病性奠定了基础。
| [1] |
Leister D. Chloroplast research in the genomic age. Trends Genet, 2003, 19: 47-56.
pmid: 12493248 |
| [2] |
Zheng Y H, Xu J M, Wang F J, Tang Y C, Wei Z, Ji Z Y, Wang C L, Zhao K J. Mutation types of cyp71p1 cause different phenotypes of mosaic spot lesion and premature leaf senescence in rice. Front Plant Sci, 2021, 12: 641300.
doi: 10.3389/fpls.2021.641300 |
| [3] |
Schippers J H, Schmidt R, Wagstaff C, Jing H C. Living to die and dying to live: the survival strategy behind leaf senescence. Plant Physiol, 2015, 169: 914-930.
doi: 10.1104/pp.15.00498 pmid: 26276844 |
| [4] |
Wingler A. Transitioning to the next phase: the role of sugar signaling throughout the plant life cycle. Plant Physiol, 2018, 176: 1075-1084.
doi: 10.1104/pp.17.01229 pmid: 28974627 |
| [5] |
Dai N, Schaffer A, Petreikov M, Shahak Y, Giller Y, Ratner K, Levine A, Granot D. Overexpression of Arabidopsis hexokinase in tomato plants inhibits growth, reduces photosynthesis, and induces rapid senescence. Plant Cell, 1999, 11: 1253-1266.
pmid: 10402427 |
| [6] |
Quirino B F, Noh Y S, Himelblau E, Amasino R M. Molecular aspects of leaf senescence. Trends Plant Sci, 2000, 5: 278-282.
pmid: 10871899 |
| [7] |
Wingler A, Delatte T L, O’Hara L E, Primavesi L F, Jhurreea D, Paul M J, Schluepmann H. Trehalose 6-phosphate is required for the onset of leaf senescence associated with high carbon availability. Plant Physiol, 2012, 158: 1241-1251.
doi: 10.1104/pp.111.191908 pmid: 22247267 |
| [8] |
Baena-González E, Sheen J. Convergent energy and stress signaling. Trends Plant Sci, 2008, 13: 474-482.
doi: 10.1016/j.tplants.2008.06.006 pmid: 18701338 |
| [9] |
Liang C, Hirose T, Okamura M, Tanimoto R, Miyao A, Hirochika H, Terao T, Li T, Ohsugi R, Aoki N. Phenotypic analyses of rice lse2 and lse3 mutants that exhibit hyperaccumulation of starch in the leaf blades. Rice, 2014, 7: 32.
doi: 10.1186/s12284-014-0032-3 |
| [10] |
肖艳华, 何光华, 陈新龙, 杜丹, 邢亚迪, 张天泉, 祝毛迪, 刘明明, 朱小燕, 桑贤春. 水稻叶片淀粉积累及早衰突变体esl9的鉴定与基因定位. 作物学报, 2017, 43: 473-482.
doi: 0.3724/SP.J.1006.2017.00473 |
| Xiao Y H, He G H, Chen X L, Du D, Xing Y D, Zhang T Q, Zhu M D, Liu M M, Zhu X Y, Sang X C. Identification and gene mapping of starch accumulation and early senescence leaf mutant esl9 in rice. Acta Agron Sin, 2017, 43: 473-482 (in Chinese with English abstract). | |
| [11] |
Chen X L, Zhu M D, Gu F X, Liu M M, Zhang Y Y, Xing Y D, Du D, Xiao Y H, Zhu X Y, He G H. Identification and gene fine mapping of starch accumulation and early senescent leaf mutant esl10 in rice. Crop Sci, 2018, 58: 204-217.
doi: 10.2135/cropsci2017.05.0323 |
| [12] |
Huang J, Yan M, Zhu X, Zhang T, Shen W Q, Yu P, Wang Y, Sang X, Yu G, Zhao B. Gene mapping of starch accumulation and premature leaf senescence in the ossac3 mutant of rice. Euphytica, 2018, 214: 177.
doi: 10.1007/s10681-018-2261-9 |
| [13] | 林添资, 孙立亭, 龚红兵, 刘玲珑, 赵志刚, 董慧, 王益华, 江玲, 万建民. 水稻叶片淀粉累积早衰突变体pls5的鉴定及基因定位. 南京农业大学学报, 2020, 43: 414-422. |
| Lin T Z, Sun L T, Gong H B, Liu L L, Zhao Z G, Dong H, Wang Y H, Jiang L, Wan J M. Identification and gene mapping of a premature leaf senescence 5 mutant with starch accumulation in rice leaves. J Nanjing Agric Univ, 2020, 43: 414-422 (in Chinese with English abstract). | |
| [14] |
Zhu M D, Chen X L, Zhu X Y, Xing Y D, Zhang T Q. Identification and gene mapping of the starch accumulation and premature leaf senescence mutant ossac4 in rice. J Integr Agric, 2020, 19: 2150-2164.
doi: 10.1016/S2095-3119(19)62814-5 |
| [15] |
Arnon D I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in beta vulgaris. Plant Physiol, 1949, 24: 1-15.
doi: 10.1104/pp.24.1.1 pmid: 16654194 |
| [16] | 邹琦. 植物生理学实验指导. 北京: 中国农业出版社, 2003. pp 160-161. |
| Zou Q. Plant Physiology Experiment. Beijing: China Agriculture Press, 2003. pp 160-161(in Chinese). | |
| [17] |
Rogers S O, Bendich A J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol Biol, 1985, 5: 69-76.
doi: 10.1007/BF00020088 pmid: 24306565 |
| [18] |
Takagi H, Uemura A, Yaegashi H, Tamiru M, Abe A, Mitsuoka C, Utsushi H, Natsume S, Kanzaki H, Matsumura H. Mutmap-gap: whole-genome resequencing of mutant f2 progeny bulk combined with de novo assembly of gap regions identifies the rice blast resistance gene pii. New Phytol, 2013, 200: 276-283.
doi: 10.1111/nph.2013.200.issue-1 |
| [19] |
Zheng C K, Zhou J J, Zhang F, Yin J J, Zhou G H, Li Y P, Chen F, Xie X Z. OsABAR1, a novel GRAM domain-containing protein, confers drought and salt tolerance via an ABA-dependent pathway in rice. Plant Physiol Biochem, 2020, 152: 138-146.
doi: 10.1016/j.plaphy.2020.04.032 |
| [20] | 王春连, 赵炳宇, 章琦, 赵开军, 邢全党. 水稻白叶枯病新抗源y238的鉴定及其近等基因系培育. 植物遗传资源学报, 2004, 5: 26-30. |
| Wang C L, Zhao B Y, Zhang Q, Zhao K J, Xing Q D. Identification of a new rice germplasm with resistance to bacterial blight and the breeding of a near-isogenic line. J Plant Genet Resour, 2004, 5: 26-30 (in Chinese with English abstract). | |
| [21] |
Barratt D H, Kölling K, Graf A, Pike M, Calder G, Findlay K, Zeeman S C, Smith A M. Callose synthase GSL7 is necessary for normal phloem transport and inflorescence growth in Arabidopsis. Plant Physiol, 2011, 155: 328-341.
doi: 10.1104/pp.110.166330 pmid: 21098675 |
| [22] |
Xie B, Wang X, Zhu M, Zhang Z, Hong Z. Cals7 encodes a callose synthase responsible for callose deposition in the phloem. Plant J, 2011, 65: 1-14.
doi: 10.1111/j.1365-313X.2010.04399.x |
| [23] |
Moghaddam M R B, Van den Ende W. Sugars and plant innate immunity. J Exp Bot, 2012, 63: 3989-3998.
doi: 10.1093/jxb/ers129 pmid: 22553288 |
| [24] | 何丹, 姜鸿瑞, 叶亚峰, 杨阳, 任艳, 陶亮之, 吴跃进, 刘斌美. 水稻早衰快腐突变体ad1衰老特性的研究. 核农学报, 2021, 35: 769-779. |
| He D, Jiang H R, Ye Y F, Yang Y, Ren Y, Tao L Z, Wu Y J, Liu B M. Analysis of the mechanism of early senescence degeneration in rice mutant ad1. J Nucl Agric Sci, 2021, 35: 769-779. | |
| [25] |
Hong Z, Zhang Z, Olson J M, Verma D P. A novel UDP-glucose transferase is part of the callose synthase complex and interacts with phragmoplastin at the forming cell plate. Plant Cell, 2001, 13: 769-779.
pmid: 11283335 |
| [26] |
Vatén A, Dettmer J, Wu S, Stierhof Y D, Miyashima S, Yadav S R, Roberts C J, Campilho A, Bulone V, Lichtenberger R, Lehesranta S, Mähönen A P, Kim J Y, Jokitalo E, Sauer N, Scheres B, Nakajima K, Carlsbecker A, Kimberly L G, Helariutta Y. Callose biosynthesis regulates symplastic trafficking during root development. Dev Cell, 2011, 21: 1144-1155.
doi: 10.1016/j.devcel.2011.10.006 pmid: 22172675 |
| [27] |
Dong X, Hong Z, Chatterjee J, Kim S, Verma D P. Expression of callose synthase genes and its connection with Npr1 signaling pathway during pathogen infection. Planta, 2008, 229: 87-98.
doi: 10.1007/s00425-008-0812-3 |
| [28] |
Xie B, Wang X, Hong Z. Precocious pollen germination in Arabidopsis plants with altered callose deposition during microsporogenesis. Planta, 2010, 231: 809-823.
doi: 10.1007/s00425-009-1091-3 pmid: 20039178 |
| [29] |
Huang L, Chen X Y, Rim Y, Han X, Cho W K, Kim S W, Kim J Y. Arabidopsis glucan synthase-like 10 functions in male gametogenesis. J Plant Physiol, 2009, 166: 344-352.
doi: 10.1016/j.jplph.2008.06.010 |
| [30] |
Nishikawa S, Zinkl G M, Swanson R J, Maruyama D, Preuss D. Callose (beta-1,3 glucan) is essential for Arabidopsis pollen wall patterning, but not tube growth. BMC Plant Biol, 2005, 5: 22.
doi: 10.1186/1471-2229-5-22 |
| [31] |
Töller A, Brownfield L, Neu C, Twell D, Schulze-Lefert P. Dual function of Arabidopsis glucan synthase-like genes GSL8 and GSL10 in male gametophyte development and plant growth. Plant J, 2008, 54: 911-923.
doi: 10.1111/j.1365-313X.2008.03462.x |
| [32] |
Dong X, Hong Z, Sivaramakrishnan M, Mahfouz M, Verma D P. Callose synthase (Cals5) is required for exine formation during microgametogenesis and for pollen viability in Arabidopsis. Plant J, 2005, 42: 315-328.
doi: 10.1111/tpj.2005.42.issue-3 |
| [33] |
Enns L C, Kanaoka M M, Torii K U, Comai L, Okada K, Cleland R E. Two callose synthases, GSL1 and GSL5, play an essential and redundant role in plant and pollen development and in fertility. Plant Mol Biol, 2005, 58: 333-349.
doi: 10.1007/s11103-005-4526-7 |
| [34] |
Jacobs A K, Lipka V, Burton R A, Panstruga R, Strizhov N, Schulze-Lefert P, Fincher G B. An Arabidopsis callose synthase, is required for wound and papillary callose formation. Plant Cell, 2003, 15: 2503-2513.
doi: 10.1105/tpc.016097 |
| [35] |
Shi X, Sun X H, Zhang Z G, Feng D, Zhang Q, Han L D, Wu J X, Lu T G. Glucan synthase-like 5 (GSL5) plays an essential role in male fertility by regulating callose metabolism during microsporogenesis in rice. Plant Cell Physiol, 2015, 56: 497-509.
doi: 10.1093/pcp/pcu193 |
| [36] | Song L Z, Wang R C, Zhang L, Wang Y M, Yao S G. CRR1 encoding callose synthase functions in ovary expansion by affecting vascular cell patterning in rice. Plant Physiol, 2016, 88: 620-632. |
| [1] | 田甜, 陈丽娟, 何华勤. 基于Meta-QTL和RNA-seq的整合分析挖掘水稻抗稻瘟病候选基因[J]. 作物学报, 2022, 48(6): 1372-1388. |
| [2] | 周文期, 强晓霞, 王森, 江静雯, 卫万荣. 水稻OsLPL2/PIR基因抗旱耐盐机制研究[J]. 作物学报, 2022, 48(6): 1401-1415. |
| [3] | 郑小龙, 周菁清, 白杨, 邵雅芳, 章林平, 胡培松, 魏祥进. 粳稻不同穗部籽粒的淀粉与垩白品质差异及分子机制[J]. 作物学报, 2022, 48(6): 1425-1436. |
| [4] | 颜佳倩, 顾逸彪, 薛张逸, 周天阳, 葛芊芊, 张耗, 刘立军, 王志琴, 顾骏飞, 杨建昌, 周振玲, 徐大勇. 耐盐性不同水稻品种对盐胁迫的响应差异及其机制[J]. 作物学报, 2022, 48(6): 1463-1475. |
| [5] | 杨建昌, 李超卿, 江贻. 稻米氨基酸含量和组分及其调控[J]. 作物学报, 2022, 48(5): 1037-1050. |
| [6] | 杨德卫, 王勋, 郑星星, 项信权, 崔海涛, 李生平, 唐定中. OsSAMS1在水稻稻瘟病抗性中的功能研究[J]. 作物学报, 2022, 48(5): 1119-1128. |
| [7] | 朱峥, 王田幸子, 陈悦, 刘玉晴, 燕高伟, 徐珊, 马金姣, 窦世娟, 李莉云, 刘国振. 水稻转录因子WRKY68在Xa21介导的抗白叶枯病反应中发挥正调控作用[J]. 作物学报, 2022, 48(5): 1129-1140. |
| [8] | 王小雷, 李炜星, 欧阳林娟, 徐杰, 陈小荣, 边建民, 胡丽芳, 彭小松, 贺晓鹏, 傅军如, 周大虎, 贺浩华, 孙晓棠, 朱昌兰. 基于染色体片段置换系群体检测水稻株型性状QTL[J]. 作物学报, 2022, 48(5): 1141-1151. |
| [9] | 王泽, 周钦阳, 刘聪, 穆悦, 郭威, 丁艳锋, 二宫正士. 基于无人机和地面图像的田间水稻冠层参数估测与评价[J]. 作物学报, 2022, 48(5): 1248-1261. |
| [10] | 陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展[J]. 作物学报, 2022, 48(4): 781-790. |
| [11] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [12] | 王吕, 崔月贞, 吴玉红, 郝兴顺, 张春辉, 王俊义, 刘怡欣, 李小刚, 秦宇航. 绿肥稻秆协同还田下氮肥减量的增产和培肥短期效应[J]. 作物学报, 2022, 48(4): 952-961. |
| [13] | 巫燕飞, 胡琴, 周棋, 杜雪竹, 盛锋. 水稻延伸因子复合体家族基因鉴定及非生物胁迫诱导表达模式分析[J]. 作物学报, 2022, 48(3): 644-655. |
| [14] | 陈云, 李思宇, 朱安, 刘昆, 张亚军, 张耗, 顾骏飞, 张伟杨, 刘立军, 杨建昌. 播种量和穗肥施氮量对优质食味直播水稻产量和品质的影响[J]. 作物学报, 2022, 48(3): 656-666. |
| [15] | 王琰, 陈志雄, 姜大刚, 张灿奎, 查满荣. 增强叶片氮素输出对水稻分蘖和碳代谢的影响[J]. 作物学报, 2022, 48(3): 739-746. |
|
||