欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (12): 3004-3017.doi: 10.3724/SP.J.1006.2022.13060
悦曼芳1,2( ), 张春2(), 郑登俞2, 邹华文1(), 吴忠义2()
), 张春2(), 郑登俞2, 邹华文1(), 吴忠义2()
YUE Man-Fang1,2(), ZHANG Chun2(), ZHENG Deng-Yu2, ZOU Hua-Wen1(), WU Zhong-Yi2()
摘要:
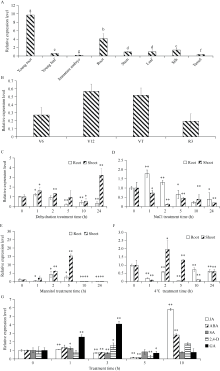
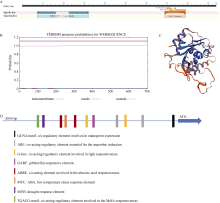
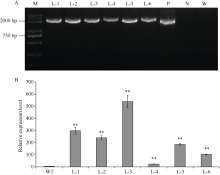
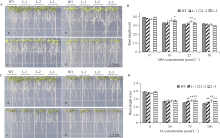
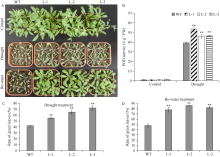
bHLH (basic helix-loop-helix)是植物中一个重要的转录因子家族, 在调控植物生长发育、逆境胁迫及信号转导过程中发挥着重要作用。目前, 动物中大部分bHLH转录因子功能已明确, 但是在植物中, 尤其是玉米中的研究报道较少。在前期工作中, 我们对玉米生长发育过程中的4个关键时期进行了根系表型鉴定和转录组测序分析, 发现转录因子ZmbHLH91在六叶期(V6)、十二叶期(V12)和抽雄期(VT)的相邻时期间的表达均差异显著, 且ZmbHLH91在V6, V12和VT这些根系生长发育活跃期的表达量较高, 推测该基因可能调控玉米根系的生长发育。为探究ZmbHLH91基因在根系生长和抵抗逆境胁迫方面的作用, 本研究克隆了ZmbHLH91 (AC: NC_AQL05369)基因, 该基因全长2112 bp, 具有bHLH转录因子家族特有的保守结构域。实时荧光定量PCR (RT-qPCR)分析表明, ZmbHLH91在玉米根中的表达量最高, 其在三叶期幼根中的表达量高于抽雄期成熟根。在高盐、渗透、低温以及脱水胁迫处理下, 玉米幼苗中ZmbHLH91的表达均上调。在无胁迫处理的1/2 MS培养基上, ZmbHLH91异源表达拟南芥与野生型拟南芥的根长无明显差别, 而在梯度浓度NaCl和甘露醇处理的培养基上, ZmbHLH91异源表达拟南芥的根均长于野生型, 且差异显著; 在土壤中进行干旱和高盐处理后, 转基因拟南芥比野生型拟南芥表现出更好的生长状态、更高的过氧化物酶(POD)活性和更高的绿叶率。推测ZmbHLH91基因可能参与响应高盐、干旱以及渗透胁迫等逆境条件。ZmbHLH91基因在茉莉酸(jasmonic acid, JA)、脱落酸(abscisic acid, ABA)等激素处理下均上调表达, 推测ZmbHLH91基因可能参与响应JA和ABA激素信号。在梯度浓度JA处理的培养基上, 转基因拟南芥的根长均长于野生型, 且差异显著。酵母双杂交实验表明ZmbHLH91与ZmMYC2相互作用, ZmMYC2是JA信号通路中重要的蛋白, 由此推测ZmbHLH91蛋白可能参与JA信号通路。综上所述, ZmbHLH91可能参与高盐、干旱和渗透胁迫应答及JA信号转导途径。本研究为进一步解析ZmbHLH91在玉米中的生物学功能提供了重要的参考依据。
| [1] |
Mao H, Yu L, Han R, Li Z, Liu H. ZmNAC55, a maize stress-responsive NAC transcription factor, confers drought resistance in transgenic Arabidopsis. Plant Physiol Biochem, 2016, 105: 55-66.
doi: 10.1016/j.plaphy.2016.04.018 |
| [2] |
Li F, Liu W. Genome-wide identification, classification, and functional analysis of the basic helix-loop-helix transcription factors in the cattle, Bos Taurus. Mamm Genome, 2017, 28: 176-197.
doi: 10.1007/s00335-017-9683-x pmid: 28299435 |
| [3] |
Murre C, McCaw P S, Vaessin H, Caudy M, Jan L Y, Jan Y N, Cabrera C V, Buskin J N, Hauschka S D, Lassar A B. Interactions between heterologous helix-loop-helix proteins generate complexes that bind specifically to a common DNA sequence. Cell, 1989, 58: 537-544.
pmid: 2503252 |
| [4] |
Jones S. An overview of the basic helix-loop-helix proteins. Genome Biol, 2004, 5: 226.
pmid: 15186484 |
| [5] |
Murre C, McCaw P S, Baltimore D. A new DNA binding and dimerization motif in immunoglobulin enhancer binding, daughterless, MyoD, and myc proteins. Cell, 1989, 56: 777-783.
pmid: 2493990 |
| [6] |
Wang C X, Qi C Y, Luo J H, Liu L, He Y, Chen L Q. Characterization of LRL5 as a key regulator of root hair growth in maize. Plant J, 2019, 98: 71-82.
doi: 10.1111/tpj.14200 |
| [7] |
Jaqueth J S, Hou Z, Zheng P, Ren R, Nagel B A, Cutter G, Niu X, Vollbrecht E, Greene T W, Kumpatla S P. Fertility restoration of maize CMS-C altered by a single amino acid substitution within the Rf4 bHLH transcription factor. Plant J, 2020, 101: 101-111.
doi: 10.1111/tpj.14521 |
| [8] |
Aslam M, Jakada B H, Fakher B, Greaves J G, Niu X, Su Z, Cheng Y, Cao S, Wang X, Qin Y. Genome-wide study of pineapple (Ananas comosus L.) bHLH transcription factors indicates that cryptochrome-interacting bHLH2 (AcCIB2) participates in flowering time regulation and abiotic stress response. BMC Genomics, 2020, 21: 735.
doi: 10.1186/s12864-020-07152-2 |
| [9] |
Fan Y, Peng J, Wu J, Zhou P, He R, Allan A C, Zeng L. NtbHLH1, a JAF13-like bHLH, interacts with NtMYB6 to enhance proanthocyanidin accumulation in Chinese Narcissus. BMC Plant Biol, 2021, 21: 275.
doi: 10.1186/s12870-021-03050-1 |
| [10] |
Guo J, Li W, Shang L, Wang Y, Yan P, Bai Y, Da X, Wang K, Guo Q, Jiang R, Mao C, Mo X. OsbHLH98 regulates leaf angle in rice through transcriptional repression of OsBUL1. New Phytol, 2021, 230: 1953-1966.
doi: 10.1111/nph.17303 |
| [11] |
Nowak K, Gaj M D. Stress-related function of bHLH109 in somatic embryo induction in Arabidopsis. J Plant Physiol, 2016, 193: 119-126.
doi: 10.1016/j.jplph.2016.02.012 |
| [12] |
Zhang L, Kang J, Xie Q, Gong J, Shen H, Chen Y, Chen G, Hu Z. The basic helix-loop-helix transcription factor bHLH95 affects fruit ripening and multiple metabolisms in tomato. J Exp Bot, 2020, 71: 6311-6327.
doi: 10.1093/jxb/eraa363 |
| [13] | Ortolan F, Fonini L S, Pastori T, Mariath J E A, Saibo N J M, Margis-Pinheiro M, Lazzarotto F. Tightly controlled expression of OsbHLH35 is critical for another development in rice. Plant Sci, 2021, 302: 110716. |
| [14] |
汪德州, 莫小婷, 张霞, 徐妙云, 张兰, 赵军, 王磊. 玉米转录因子ZmbHLH4基因的克隆及功能分析. 中国农业科技导报, 2018, 20(12): 16-25.
doi: 10.13304/j.nykjdb.2018.0189 |
| Wang D Z, Mo X T, Zhang X, Xu M Y, Zhang L, Zhao J, Wang L. Cloning and functional analysis of transcription factors gene ZmbHLH4 from Zea mays. J Agric Sci Technol, 2018, 20(12): 16-25. (in Chinese with English abstract) | |
| [15] |
Yang Y Y, Zheng P F, Ren Y R, Yao Y X, You C X, Wang X F, Hao Y J. Apple MdSAT1 encodes a bHLHm1 transcription factor involved in salinity and drought responses. Planta, 2021, 253: 46.
doi: 10.1007/s00425-020-03528-6 |
| [16] |
Liu Y, Ji X, Nie X, Qu M, Zheng L, Tan Z, Zhao H, Huo L, Liu S, Zhang B, Wang Y. Arabidopsis AtbHLH112 regulates the expression of genes involved in abiotic stress tolerance by binding to their E-box and GCG-box motifs. New Phytol, 2015, 207: 692-709.
doi: 10.1111/nph.13387 |
| [17] |
Yang X, Wang R, Hu Q, Li S, Mao X, Jing H, Zhao J, Hu G, Fu J, Liu C. DlICE1, a stress-responsive gene from Dimocarpus longan, enhances cold tolerance in transgenic Arabidopsis. Plant Physiol Biochem, 2019, 142: 490-499.
doi: 10.1016/j.plaphy.2019.08.007 |
| [18] | Zhao Q, Fan Z, Qiu L, Che Q, Wang T, Li Y, Wang Y. MdbHLH130, an apple bHLH transcription factor, confers water stress resistance by regulating stomatal closure and ROS homeostasis in transgenic tobacco. Front Plant Sci, 2020, 11: 543696. |
| [19] | Babitha K C, Vemanna R S, Nataraja K N, Udayakumar M. Overexpression of EcbHLH57 transcription factor from Eleusine coracana L. in tobacco confers tolerance to salt, oxidative and drought stress. PLoS One, 2015, 10: e137098. |
| [20] | Qian Y, Zhang T, Yu Y, Gou L, Yang J, Xu J, Pi E. Regulatory mechanisms of bHLH transcription factors in plant adaptive responses to various abiotic stresses. Front Plant Sci, 2021, 12: 677611. |
| [21] |
Li Z, Liu C, Zhang Y, Wang B, Ran Q, Zhang J. The bHLH family member ZmPTF1 regulates drought tolerance in maize by promoting root development and abscisic acid synthesis. J Exp Bot, 2019, 70: 5471-5486.
doi: 10.1093/jxb/erz307 |
| [22] | Chen H C, Cheng W H, Hong C Y, Chang Y S, Chang M C. The transcription factor OsbHLH035 mediates seed germination and enables seedling recovery from salt stress through ABA- dependent and ABA-independent pathways, respectively. Rice (New York), 2018, 11: 1-17. |
| [23] |
Zhao Q, Xiang X, Liu D, Yang A, Wang Y. Tobacco transcription factor NtbHLH123 confers tolerance to cold stress by regulating the NtCBF pathway and reactive oxygen species homeostasis. Front Plant Sci, 2018, 9: 381.
doi: 10.3389/fpls.2018.00381 |
| [24] |
Liu H, Yang Y, Liu D, Wang X, Zhang L. Transcription factor TabHLH49 positively regulates dehydrin WZY2gene expression and enhances drought stress tolerance in wheat. BMC Plant Biol, 2020, 20: 259.
doi: 10.1186/s12870-020-02474-5 |
| [25] |
Wei S, Xia R, Chen C, Shang X, Ge F, Wei H, Chen H, Wu Y, Xie Q. ZmbHLH124 identified in maize recombinant inbred lines contributes to drought tolerance in crops. Plant Biotechnol J, 2021, 19: 2069-2081.
doi: 10.1111/pbi.13637 |
| [26] |
杨梦婷, 张春, 王作平, 邹华文, 吴忠义. 玉米ZmbHLH161基因的克隆及功能研究. 作物学报, 2020, 46: 2008-2016.
doi: 10.3724/SP.J.1006.2020.03022 |
| Yang M T, Zhang C, Wang Z P, Zou H W, Wu Z Y. Cloning and functional analysis of ZmbHLH161 gene in maize. Acta Agron Sin, 2020, 46: 2008-2016. (in Chinese with English abstract) | |
| [27] |
Rabissi A, Vilela B, Lumbreras V, Ludevid D, Culiáñez-Macià F A, Pagés M. Molecular characterization of maize bHLH transcription factor (ZmKS), a new ZmOST1 kinase substrate. Plant Sci, 2016, 253: 1-12.
doi: S0168-9452(16)30423-X pmid: 27968978 |
| [28] |
Duek P D, Fankhauser C. HFR1, a putative bHLH transcription factor, mediates both phytochrome A and cryptochrome signalling. Plant J, 2003, 34: 827-836.
pmid: 12795702 |
| [29] |
Kiribuchi K, Jikumaru Y, Kaku H, Minami E, Hasegawa M, Kodama O, Seto H, Okada K, Nojiri H, Yamane H. Involvement of the basic helix-loop-helix transcription factor RERJ1in wounding and drought stress responses in rice plants. Biosci Biotechnol Biochem, 2005, 69: 1042-1044.
doi: 10.1271/bbb.69.1042 |
| [30] |
Castillon A, Shen H, Huq E. Phytochrome Interacting Factors: central players in phytochrome-mediated light signaling networks. Trends Plant Sci, 2007, 12: 514-521.
doi: 10.1016/j.tplants.2007.10.001 pmid: 17933576 |
| [31] |
Friedrichsen D M, Nemhauser J, Muramitsu T, Maloof J N, Alonso J, Ecker J R, Furuya M, Chory J. Three redundant brassinosteroid early response genes encode putative bHLH transcription factors required for normal growth. Genetics, 2002, 162: 1445-1456.
doi: 10.1093/genetics/162.3.1445 pmid: 12454087 |
| [32] |
Seo J S, Joo J, Kim M J, Kim Y K, Nahm B H, Song S I, Cheong J J, Lee J S, Kim J K, Choi Y D. OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J, 2011, 65: 907-921.
doi: 10.1111/j.1365-313X.2010.04477.x |
| [33] |
Zhang T, Lv W, Zhang H, Ma L, Li P, Ge L, Li G. Genome-wide analysis of the basic Helix-Loop-Helix (bHLH) transcription factor family in maize. BMC Plant Biol, 2018, 18: 235.
doi: 10.1186/s12870-018-1441-z pmid: 30326829 |
| [34] |
Zhang C, Li X, Wang Z, Zhang Z, Wu Z. Identifying key regulatory genes of maize root growth and development by RNA sequencing. Genomics, 2020, 112: 5157-5169.
doi: 10.1016/j.ygeno.2020.09.030 pmid: 32961281 |
| [35] |
Clough S J, Bent A F. Floral dip: a simplified method for Agrobacterium mediated transformation of Arabidopsis thaliana. Plant J, 1998, 16: 735-743.
pmid: 10069079 |
| [36] |
Chini A, Fonseca S, Fernández G, Adie B, Chico J M, Lorenzo O, García-Casado G, López-Vidriero I, Lozano F M, Ponce M R, Micol J L, Solano R. The JAZ family of repressors is the missing link in jasmonate signalling. Nature, 2007, 448: 666-671.
doi: 10.1038/nature06006 |
| [37] |
Fernández-Calvo P, Chini A, Fernández-Barbero G, Chico J M, Gimenez-Ibanez S, Geerinck J, Eeckhout D, Schweizer F, Godoy M, Franco-Zorrilla J M, Pauwels L, Witters E, Puga M I, Paz-Ares J, Goossens A, Reymond P, De Jaeger G, Solano R. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell, 2011, 23: 701-715.
doi: 10.1105/tpc.110.080788 |
| [38] |
Liu X, Galli M, Camehl I, Gallavotti A. RAMOSA1 ENHANCER LOCUS2-mediated transcriptional repression regulates vegetative and reproductive architecture. Plant Physiol, 2019, 179: 348-363.
doi: 10.1104/pp.18.00913 pmid: 30348817 |
| [39] |
Fu J, Liu L, Liu Q, Shen Q, Wang C, Yang P, Zhu C, Wang Q. ZmMYC2 exhibits diverse functions and enhances JA signaling in transgenic Arabidopsis. Plant Cell Rep, 2020, 39: 273-288.
doi: 10.1007/s00299-019-02490-2 |
| [1] | 孙智超, 张吉旺. 弱光胁迫影响玉米产量形成的生理机制及调控效应[J]. 作物学报, 2023, 49(1): 12-23. |
| [2] | 濮雪, 王凯彤, 张宁, 司怀军. 马铃薯StMAPKK4基因表达分析及互作蛋白筛选与鉴定[J]. 作物学报, 2023, 49(1): 36-45. |
| [3] | 陈冰洁, 张富粮, 杨硕, 李晓立, 何堂庆, 张晨曦, 田明慧, 吴梅, 郝晓峰, 张学林. 不同形态氮肥下丛枝菌根真菌对玉米灌浆期生理特性及产量和品质的影响[J]. 作物学报, 2023, 49(1): 249-261. |
| [4] | 张静, 王洪章, 任昊, 殷复伟, 吴红燕, 赵斌, 张吉旺, 任佰朝, 戴爱斌, 刘鹏. 夏玉米根系构型与抗根倒性能间的关系[J]. 作物学报, 2023, 49(1): 188-199. |
| [5] | 王锐璞, 董振营, 高悦欣, 鲍建喜, 殷芳冰, 李金萍, 龙艳, 万向元. 玉米籽粒淀粉含量全基因组关联分析和候选基因预测[J]. 作物学报, 2023, 49(1): 140-152. |
| [6] | 商蒙非, 石晓宇, 赵炯超, 李硕, 褚庆全. 气候变化背景下中国不同区域玉米生育期高温胁迫时空变化特征[J]. 作物学报, 2023, 49(1): 167-176. |
| [7] | 段灿星, 崔丽娜, 夏玉生, 董怀玉, 杨知还, 胡清玉, 孙素丽, 李晓, 朱振东, 王晓鸣. 玉米种质资源对拟轮枝镰孢与禾谷镰孢穗腐病的抗性精准鉴定与分析[J]. 作物学报, 2022, 48(9): 2155-2167. |
| [8] | 张振博, 屈馨月, 于宁宁, 任佰朝, 刘鹏, 赵斌, 张吉旺. 施氮量对夏玉米籽粒灌浆特性和内源激素作用的影响[J]. 作物学报, 2022, 48(9): 2366-2376. |
| [9] | 郭瑶, 柴强, 殷文, 范虹. 玉米密植光合生理机制及应用途径研究进展[J]. 作物学报, 2022, 48(8): 1871-1883. |
| [10] | 王天波, 赫文学, 张峻铭, 吕伟增, 梁雨欢, 卢洋, 王雨露, 谷丰序, 宋词, 陈军营. 人工老化玉米种胚ROS产生及ATP合成酶亚基mRNA完整性研究[J]. 作物学报, 2022, 48(8): 1996-2006. |
| [11] | 裴丽珍, 陈远学, 张雯雯, 肖华, 张森, 周元, 徐开未. 有机物料还田对夏玉米穗位叶光合性能及氮代谢的影响[J]. 作物学报, 2022, 48(8): 2115-2124. |
| [12] | 郭家鑫, 鲁晓宇, 陶一凡, 郭慧娟, 闵伟. 棉花在盐碱胁迫下代谢产物及通路的分析[J]. 作物学报, 2022, 48(8): 2100-2114. |
| [13] | 杨迎霞, 张冠, 王梦梦, 陆国清, 王倩, 陈锐. 基于高通量测序技术的转基因玉米GM11061分子特征研究[J]. 作物学报, 2022, 48(7): 1843-1850. |
| [14] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [15] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
|