欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (10): 2705-2716.doi: 10.3724/SP.J.1006.2023.24261
朱继杰1( ), 王士杰1, 赵红霞1, 贾晓昀1(), 李妙1(), 王国印2
), 王士杰1, 赵红霞1, 贾晓昀1(), 李妙1(), 王国印2
ZHU Ji-Jie1(), WANG Shi-Jie1, ZHAO Hong-Xia1, JIA Xiao-Yun1(), LI Miao1(), WANG Guo-Yin2
摘要:
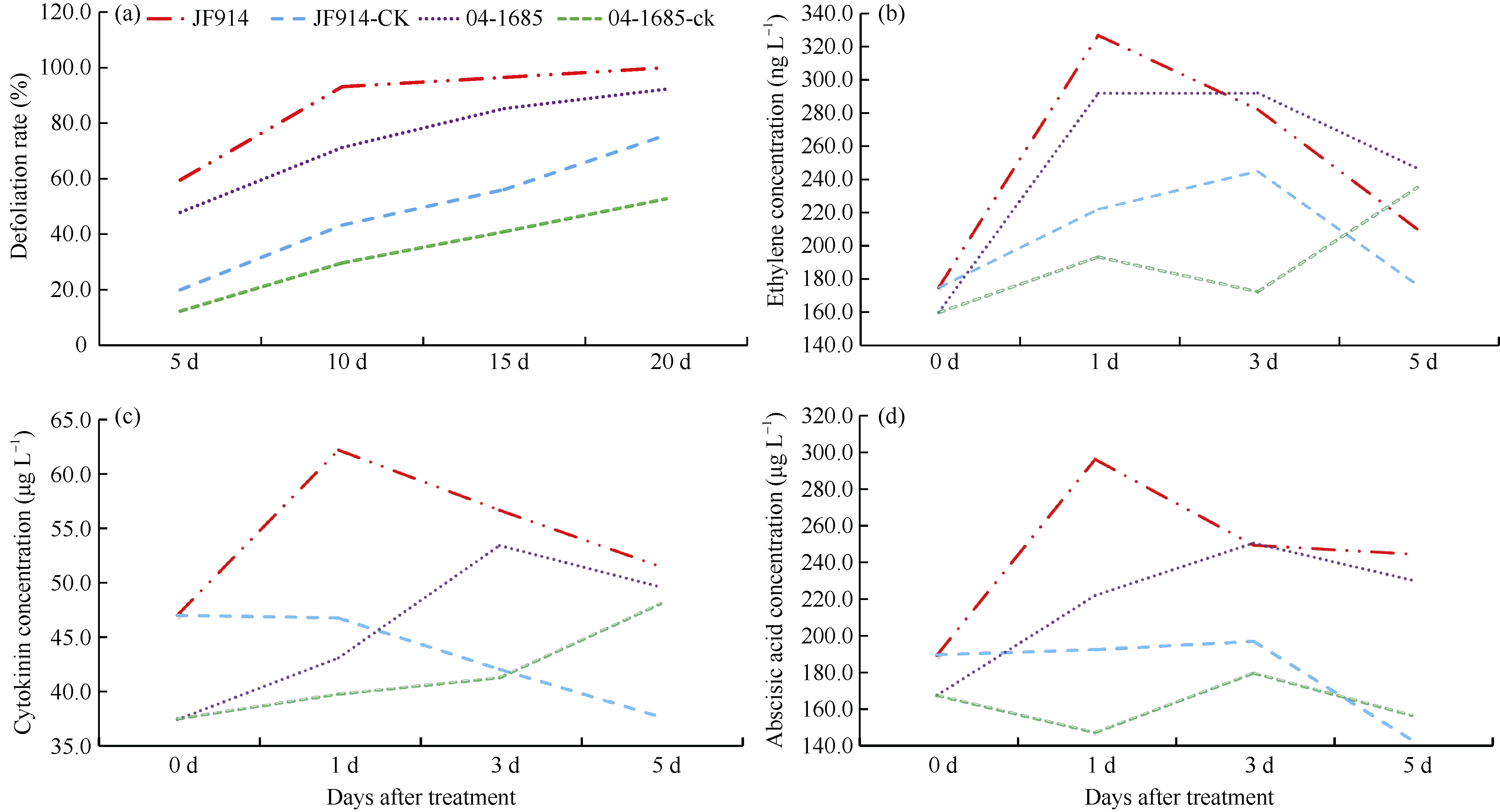
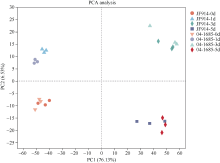
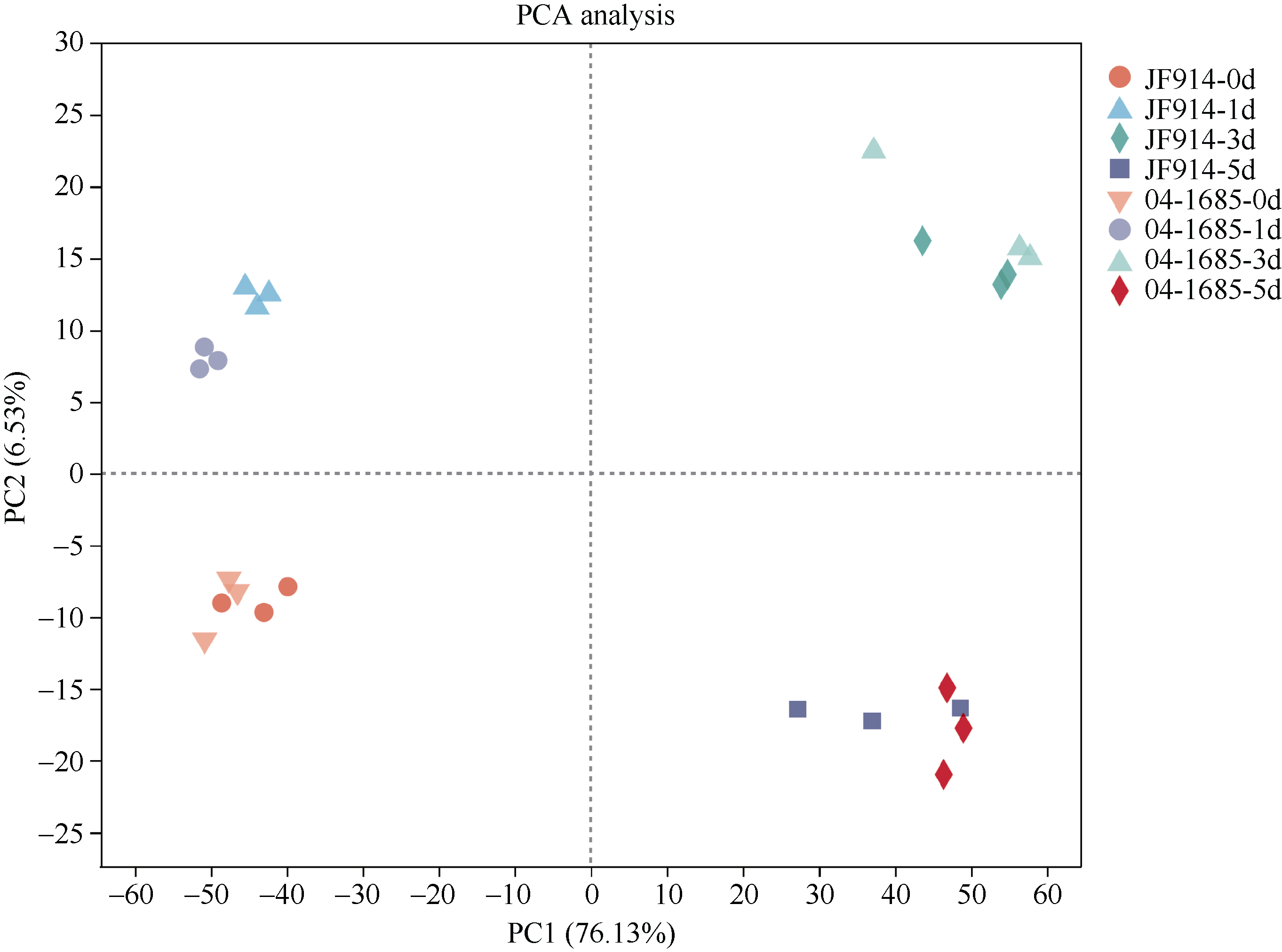
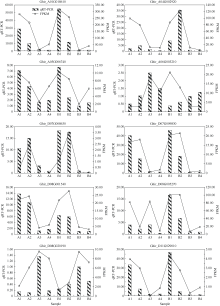
噻苯隆是棉花脱叶的主要化学药剂, 在实现棉花机械采收中发挥了关键作用。目前, 噻苯隆在大田环境下诱导棉花叶片脱落的分子调控机制研究较少。本研究以噻苯隆敏感型品种(冀丰914)和不敏感型株系(04-1685)为材料, 测定叶片的乙烯、细胞分裂素和脱落酸含量, 同时利用RNA-seq技术, 分析噻苯隆处理后0 d、1 d、3 d、5 d叶片的基因表达变化, 利用WGCNA (Weighted gene coexpression network analysis)分析筛选基因共表达模块, 利用GO和KEGG分析基因功能。结果表明, 冀丰914的乙烯、细胞分裂素和脱落酸含量在处理后1 d达到最高, 04-1685的激素含量在处理后3 d达到最高。通过RNA-seq筛选到33,283个差异表达基因(differentially expressed genes, DEGs), 通过WGCNA分析, 分别筛选到2个和3个与激素和样本高度相关的模块, 模块内的基因主要参与非生物胁迫响应、昼夜节律、物质代谢等过程。通过分析植物激素、胁迫响应、昼夜节律等主要过程相关基因的表达量发现, 激素相关基因在敏感型品种冀丰914中的表达量更高, 其中乙烯信号响应基因在处理后5 d内均保持在较高的表达水平, 脱落酸信号响应基因在处理后1 d内保持较高的表达水平, 细胞分裂素代谢相关基因在处理后1 d内表达量很低, 处理后3 d和5 d的表达量急剧增加。处理后1 d和3 d, 热激蛋白基因(HSP70、HSP90、ATJ3)、钙稳态调控基因(CRT3、NCL)和节律基因(LNK2、APRR9、CCA1)显著下调表达, 节律基因(ARR8)上调表达。大田环境下喷施噻苯隆, 在引起激素含量变化的同时, 可能破坏了植物的昼夜节律、降低对非生物胁迫的应激能力, 从而加速叶片衰老和脱落。本研究为丰富噻苯隆诱导叶片脱落的分子调控机制提供了更多思路。
| [1] |
Jin D, Xu Y, Gui H, Zhang H, Dong Q, Sikder R K, Wang X, Yang G, Song M. Evaluation of cotton (Gossypium hirsutum L.) leaf abscission sensitivity triggered by thidiazuron through membership function value. Plants, 2021, 10: 49.
doi: 10.3390/plants10010049 |
| [2] |
田景山, 张煦怡, 张丽娜, 徐守振, 祁炳琴, 随龙龙, 张鹏鹏, 杨延龙, 张旺锋, 勾玲. 新疆机采棉花实现叶片快速脱落需要的温度条件. 作物学报, 2019, 45: 613-620.
doi: 10.3724/SP.J.1006.2019.84068 |
|
Tian J S, Zhang X Y, Zhang L N, Xu S Z, Qi B Q, Sui L L, Zhang P P, Yang Y L, Zhang W F, Gou L. Temperatures of promoting rapid leaf abscission of cotton in Xinjiang region. Acta Agron Sin, 2019, 45: 613-620. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2019.84068 |
|
| [3] |
聂军军, 代建龙, 杜明伟, 张艳军, 田晓莉, 李召虎, 董合忠. 我国现代植棉理论与技术的新发展: 棉花集中成熟栽培. 中国农业科学, 2021, 54: 4286-4298.
doi: 10.3864/j.issn.0578-1752.2021.20.004 |
| Nie J J, Dai J L, Du M W, Zhang Y J, Tian X L, Li Z H, Dong H Z. New development of modern cotton farming theory and technology in China: concentrated maturation cultivation of cotton. Sci Agric Sin, 2021, 54: 4286-4298. (in Chinese with English abstract) | |
| [4] | 高丽丽, 李淦, 康正华, 李健伟, 王蜜蜂, 马云珍, 张巨松. 脱叶剂对棉花叶片叶绿素荧光动力学参数的影响. 棉花学报, 2016, 28: 345-352. |
| Gao L L, Li G, Kang Z H, Li J W, Wang M F, Ma Y Z, Zhang J S. Effect of defoliants on chlorophyll fluorescence of cotton leaves. Cotton Sci, 2016, 28: 345-352. (in Chinese with English abstract) | |
| [5] | 宋兴虎, 徐东永, 孙璐, 赵文超, 曹龙龙, 张祥, 唐纪元, 韩焕勇, 王洪这, 陈洪章, 王林, 赵冰梅, 杜明伟, 田晓莉, 李召虎. 在不同棉区噻苯隆和乙烯利用量及配比对脱叶催熟效果影响. 棉花学报, 2020, 32: 247-257. |
| Song X H, Xu D Y, Sun L, Zhao W C, Cao L L, Zhang X, Tang J Y, Han H Y, Wang H Z, Chen H Z, Wang L, Zhao B M, Du M W, Tian X L, Li Z H. Effect of thidiazuron and ethylene use and ratio on defoliation ripening in different cotton area. Cotton Sci, 2020, 32: 247-257. (in Chinese with English abstract) | |
| [6] |
张大伟, 魏鑫, 徐海江, 刘忠山, 李春平, 马清倩, 徐建辉. 不同棉花品种对脱叶剂的响应. 新疆农业科学, 2019, 56(1): 146-153.
doi: 10.6048/j.issn.1001-4330.2019.01.018 |
| Zhang D W, Wei X, Xu H J, Liu Z S, Li C P, Ma Q Q, Xu J H. Study on the response of cotton varieties with different genotypes to defoliants. Xinjiang Agric Sci, 2019, 56(1): 146-153. (in Chinese with English abstract) | |
| [7] | 王天友. 南疆陆地棉种质资源遗传多样性及对脱叶剂的敏感性分析. 塔里木大学硕士学位论文,新疆阿拉尔, 2020. |
| Wang T Y. Analysis of Genetic Diversity and Sensitivity to Sefoloant of Upland Cotton Germplasm Resources in Southern Xinjiang. MS Thesis of Tarim University, Aral, Xinjiang, China, 2020. (in Chinese with English abstract) | |
| [8] |
周先林, 覃琴, 王龙, 李璐, 胡成成, 洪秀春, 王伟, 朱海勇. 脱叶剂对两种机采模式下棉花脱叶效果及纤维品质的影响. 中国农业科技导报, 2020, 22(11): 144-152.
doi: 10.13304/j.nykjdb.2019.0628 |
| Zhou X L, Qin Q, Wang L, Li L, Hu C C, Hong X C, Wang W, Zhu H Y. Influence of defoliant on defoliation effect and fiber quality of cotton under two kinds of mechanical harvesting modes. J Agric Sci Technol, 2020, 22(11): 144-152. (in Chinese with English abstract) | |
| [9] |
Suttle J C. Disruption of the polar auxin transport system in cotton seedlings following treatment with the defoliant thidiazuron. Plant Physiol, 1988, 86: 241-245.
doi: 10.1104/pp.86.1.241 pmid: 16665874 |
| [10] |
Li F, Wu Q, Liao B, Yu K, Huo Y, Meng L, Wang S, Wang B, Du M, Tian X, Li Z. Thidiazuron promotes leaf abscission by regulating the crosstalk complexities between ethylene, auxin, and cytokinin in cotton. Int J Mol Sci, 2022, 23: 2696.
doi: 10.3390/ijms23052696 |
| [11] | 廖宝鹏, 王崧嫚, 杜明伟, 李芳军, 田晓莉, 李召虎. 棉花不同部位主茎叶对脱叶剂噻苯隆的响应及机理. 棉花学报, 2020, 32: 418-424. |
| Liao B P, Wang S M, Du M W, Li F J, Tian X L, Li Z H. Responses and underlying mechanisms of different mainstem leaves on cotton to defoliant thidiazuron. Cotton Sci, 2020, 32: 418-424. (in Chinese with English abstract) | |
| [12] | 高瑜, 徐娇, 张冰, 孙伟男, 杨细燕. 机采棉化学脱叶伴随着剧烈的乙烯及细胞分裂素信号响应. 棉花学报, 2020, 32: 491-500. |
| Gao Y, Xu J, Zhang B, Sun W N, Yang X Y. Chemical defoliation of machine-harvested cotton was accompanied by intense ethylene and cytokinin signal responses. Cotton Sci, 2020, 32: 491-500. (in Chinese with English abstract) | |
| [13] |
Xu J, Chen L, Sun H, Wusiman N, Sun W, Li B, Gao Y, Kong J, Zhang D, Zhang X, Xu H, Yang X. Crosstalk between cytokinin and ethylene signaling pathways regulates leaf abscission in cotton in response to chemical defoliants. J Exp Bot, 2019, 70: 1525-1538.
doi: 10.1093/jxb/erz036 pmid: 30715415 |
| [14] |
Jin D, Wang X, Xu Y, Gui H, Zhang H, Dong Q, Sikder R K, Yang G, Song M. Chemical defoliant promotes leaf abscission by altering ROS metabolism and photosynthetic efficiency in Gossypium hirsutum. Int J Mol Sci, 2020, 21: 2738.
doi: 10.3390/ijms21082738 |
| [15] | 王晓婧, 李思嘉, 刘瑞显, 张国伟, 杨长琴, 倪万潮. 棉花施用脱叶剂对相邻未着药叶片生理活性的影响. 棉花学报, 2019, 31: 64-71. |
| Wang X J, Li S J, Liu R X, Zhang G W, Yang C Q, Ni W C. Effect of defoliants application on physiological characters of cotton leaf without defoliants. Cotton Sci, 2019, 31: 64-71. (in Chinese with English abstract) | |
| [16] | 高丽丽. 脱叶剂喷施时间对棉花生理调节效应的研究. 新疆农业大学硕士学位论文, 新疆乌鲁木齐, 2016. |
| Gao L L. Study of Defoliants Spraying Time on Cotton Physiological Mechanism. MS Thesis of Xinjiang Agricultural University, Urumqi, Xinjiang, China, 2016. (in Chinese with English abstract) | |
| [17] | 朱继杰, 王士杰, 赵红霞, 李妙, 王国印, 聂俊杰, 牛立强, 闫丽丽. 适宜河北省机械采摘的棉花品种筛选研究. 河北农业科学, 2018, 22(4): 60-62. |
| Zhu J J, Wang S J, Zhao H X, Li M, Wang G Y, Nie J J, Niu L L, Yan L L. Study on screening cotton varieties suitable for mechanical harvest in Hebei province. J Hebei Agric Sci, 2018, 22(4): 60-62. (in Chinese with English abstract) | |
| [18] | 朱继杰, 赵红霞, 王士杰, 李妙, 王国印. 不同棉花品种对脱叶剂敏感性研究. 中国棉花, 2018, 45(4): 15-18. |
| Zhu J J, Zhao H X, Wang S J, Li M, Wang G Y. Study on the sensitivity of different cotton cultivars to defoliant. China Cotton, 2018, 45(4): 15-18. (in Chinese with English abstract) | |
| [19] |
Xu X, Yuan L, Yang X, Zhang X, Wang L, Xie Q. Circadian clock in plants: linking timing to fitness. J Integr Plant Biol, 2022, 64: 792-811.
doi: 10.1111/jipb.13230 |
| [20] |
Creux N, Harmer S. Circadian rhythms in plants. Cold Spring Harb Perspect Biol, 2019, 11: a034611.
doi: 10.1101/cshperspect.a034611 |
| [21] |
Srivastava D, Shamim M, Kumar M, Mishra A, Maurya R, Sharma D, Pandey P, Singh K N. Role of circadian rhythm in plant system: an update from development to stress response. Environ Exp Bot, 2019, 162: 256-271.
doi: 10.1016/j.envexpbot.2019.02.025 |
| [22] |
Wahid A, Gelani S, Ashraf M, Foolad M. Heat tolerance in plants: an overview. Environ Exp Bot, 2007, 61: 199-223.
doi: 10.1016/j.envexpbot.2007.05.011 |
| [23] | Xu Y, Zhan C, Huang B. Heat Shock Proteins in association with heat tolerance in grasses. Int J Proteom, 2011, 2011: 529648. |
| [24] |
Hasanuzzaman M, Nahar K, Alam M, Roychowdhury R, Fujita M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int J Mol Sci, 2013, 14: 9643-9684.
doi: 10.3390/ijms14059643 pmid: 23644891 |
| [25] |
Hanano S, Domagalska M A, Nagy F, Davis S J. Multiple phytohormones influence distinct parameters of the plant circadian clock. Genes Cells, 2006, 11: 1381-1392.
pmid: 17121545 |
| [26] |
Covington M F, Maloof J N, Straume M, Kay S A, Harmer S L. Global transcriptome analysis reveals circadian regulation of key pathways in plant growth and development. Genom Biol, 2008, 9: R130.
doi: 10.1186/gb-2008-9-8-r130 |
| [27] |
Song Q, Ando A, Xu D, Fang L, Zhang T, Hu E, Qiao H, Deng X W, Chen Z J. Diurnal down-regulation of ethylene biosynthesis mediates biomass heterosis. Proc Natl Acad Sci USA, 2018, 115: 5606-5611.
doi: 10.1073/pnas.1722068115 pmid: 29735680 |
| [28] |
Haydon M J, Mielczarek O, Frank A, Román Á, Webb A A. Sucrose and ethylene signaling interact to modulate the circadian clock. Plant Physiol, 2017, 175: 947-958.
doi: 10.1104/pp.17.00592 pmid: 28778922 |
| [29] |
Mockler T C, Michael T P, Priest H D, Shen R, Sullivan C M, Givan S A, McEntee C, Kay S A, Chory J. The DIURNAL project: DIURNAL and circadian expression profiling, model-based pattern matching, and promoter analysis. Cold Spring Harb Symp Quant Biol, 2007, 72: 353-363.
doi: 10.1101/sqb.2007.72.006 pmid: 18419293 |
| [30] |
Kou X, Zhao X, Wu B, Wang C, Wu C, Yang S, Zhou J, Xue Z. Auxin response factors are ubiquitous in plant growth and development, and involved in crosstalk between plant hormones: a review. Appl Sci, 2022, 12: 1360.
doi: 10.3390/app12031360 |
| [31] |
Kamal N M, Gorafi Y S A, Abdelrahman M, Abdellatef E, Tsujimoto H. Stay-green trait: a prospective approach for yield potential, and drought and heat stress adaptation in globally important cereals. Int J Mol Sci, 2019, 20: 5837.
doi: 10.3390/ijms20235837 |
| [32] |
Ahanger M A, Ashraf M, Bajguz A, Ahmad P. Brassinosteroids regulate growth in plants under stressful environments and crosstalk with other potential phytohormones. J Plant Growth Regul, 2018, 37: 1007-1024.
doi: 10.1007/s00344-018-9855-2 |
| [1] | 王菲菲, 张胜忠, 胡晓辉, 崔凤高, 钟文, 赵立波, 张天雨, 郭进涛, 于豪谅, 苗华荣, 陈静. 比较转录组分析花生种子休眠调控网络[J]. 作物学报, 2023, 49(9): 2446-2461. |
| [2] | 胡鑫, 罗正英, 李纯佳, 吴转娣, 李旭娟, 刘新龙. 基于二代和三代转录组测序揭示甘蔗重要亲本对黑穗病菌侵染的响应机制[J]. 作物学报, 2023, 49(9): 2412-2432. |
| [3] | 左春阳, 李亚玮, 李焱龙, 金双侠, 朱龙付, 张献龙, 闵玲. 陆地棉漆酶基因家族成员表达模式分析[J]. 作物学报, 2023, 49(9): 2344-2361. |
| [4] | 文利超, 熊涛, 邓智超, 刘涛, 郭存, 李伟, 郭永峰. 烟草转录因子NtNAC080在非生物胁迫下的表达分析及功能鉴定[J]. 作物学报, 2023, 49(8): 2171-2182. |
| [5] | 陈力, 王靖, 邱晓, 孙海莲, 张文浩, 王天佐. 不同耐旱性紫花苜蓿干旱胁迫下生理响应和转录调控的差异研究[J]. 作物学报, 2023, 49(8): 2122-2132. |
| [6] | 丁洪艳, 冯晓溪, 汪柏宇, 张积森. 甘蔗割手密种LRRII-RLK基因家族演化和表达分析[J]. 作物学报, 2023, 49(7): 1769-1784. |
| [7] | 王会伟, 张向歌, 李春鑫, 许欣然, 胡海燕, 朱雅婧, 王艳, 张新友. 油莎豆耐盐性评估及盐胁迫下幼苗根系转录组学分析[J]. 作物学报, 2023, 49(7): 1882-1894. |
| [8] | 李凌雨, 周琦锐, 李洋, 张安民, 王贝贝, 马尚宇, 樊永惠, 黄正来, 张文静. 外源6-BA调控孕穗期低温后小麦幼穗发育的转录组分析[J]. 作物学报, 2023, 49(7): 1808-1817. |
| [9] | 王雁楠, 陈金金, 卞倩倩, 胡琳琳, 张莉, 尹雨萌, 乔守晨, 曹郭郑, 康志河, 赵国瑞, 杨国红, 杨育峰. 转录组与代谢组联合分析揭示遮阴胁迫下甘薯的代谢响应途径[J]. 作物学报, 2023, 49(7): 1785-1798. |
| [10] | 马春敏, 李维希, 李芳军, 田晓莉, 李召虎. 陆地棉硝酸盐转运体NRT基因家族鉴定及表达分析[J]. 作物学报, 2023, 49(6): 1496-1517. |
| [11] | 张小红, 彭琼, 鄢铮. 盐胁迫下不同甘薯品种的转录组测序分析[J]. 作物学报, 2023, 49(5): 1432-1444. |
| [12] | 孙全喜, 苑翠玲, 牟艺菲, 闫彩霞, 赵小波, 王娟, 王奇, 孙慧, 李春娟, 单世华. 花生SWEET基因全基因组鉴定及表达分析[J]. 作物学报, 2023, 49(4): 938-954. |
| [13] | 周宾寒, 杨竹, 王书平, 方正武, 胡赞民, 徐兆师, 张迎新. 小麦幼苗活性LTR反转录转座子筛选及其对非生物胁迫的响应[J]. 作物学报, 2023, 49(4): 966-977. |
| [14] | 徐子寅, 于晓玲, 邹良平, 赵平娟, 李文彬, 耿梦婷, 阮孟斌. 木薯MYB转录因子基因MeMYB60表达特征分析及其互作蛋白筛选[J]. 作物学报, 2023, 49(4): 955-965. |
| [15] | 郭宏, 于霁雯, 裴文锋, 关永虎, 李航, 李长喜, 刘金伟, 王伟, 王宝全, 梅拥军. 南疆陆地棉杂种F2的遗传分析及遗传主效聚类[J]. 作物学报, 2023, 49(3): 608-621. |
|