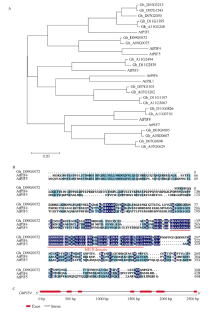
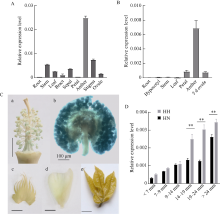
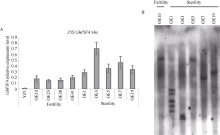
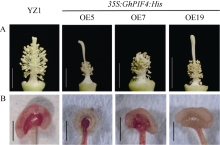
Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2020, Vol. 46 ›› Issue (9): 1368-1379.doi: 10.3724/SP.J.1006.2020.94188
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
CHEN Miao( ), XIE Sai, WANG Chao-Zhi, LI Yan-Long, ZHANG Xian-Long, MIN Ling*()
), XIE Sai, WANG Chao-Zhi, LI Yan-Long, ZHANG Xian-Long, MIN Ling*()
| [1] |
Zhang Q, Shen B Z, Dai X K, Mei M H, Saghai Maroof M A, Li Z B. Using bulked extremes and recessive class to map genes for photoperiod-sensitive genic male sterility in rice. Proc Natl Acad Sci USA, 1994,91:8675-8679.
pmid: 7915844 |
| [2] |
Chen L, Liu Y G. Male sterility and fertility restoration in crops. Annu Rev Plant Biol, 2014,65:579-606.
doi: 10.1146/annurev-arplant-050213-040119 pmid: 24313845 |
| [3] | 袁隆平. 选育水稻光、温敏核不育系的技术策略. 杂交水稻, 1992, (1):1-4. |
| Yuan L P. Technical strategy for breeding rice light and temperature sensitive genic male sterile lines. Hybrid Rice, 1992, (1):1-4 (in Chinese). | |
| [4] |
Endo M, Tsuchiya T, Hamada K, Kawamura S, Yano K, Ohshima M, Higashitani A, Watanabe M, Kawagishi-Kobayashi M. High temperatures cause male sterility in rice plants with transcriptional alterations during pollen development. Plant Cell Physiol, 2009,50:1911-1922.
doi: 10.1093/pcp/pcp135 pmid: 19808807 |
| [5] |
Zinn K E, Tunc-Ozdemir M, Harper J F. Temperature stress and plant sexual reproduction: uncovering the weakest links. J Exp Bot, 2010,61:1959-1968.
pmid: 20351019 |
| [6] |
Dong B, Zheng X, Liu H P, Able J A, Yang H, Zhao H, Zhang M M, Qiao Y Z, Wang Y K, Liu M Y. Effects of drought stress on pollen sterility, grain yield, abscisic acid and protective enzymes in two winter wheat cultivars. Front Plant Sci, 2017,8:1008.
pmid: 28676806 |
| [7] |
De Storme N, Geelen D. The impact of environmental stress on male reproductive development in plants: biological processes and molecular mechanisms. Plant Cell Environ, 2014,37:1-18.
doi: 10.1111/pce.12142 pmid: 23731015 |
| [8] |
Khanna R, Huq E, Kikis E A, Al-Sady B, Lanzatella C, Quail P H. A novel molecular recognition motif necessary for targeting photoactivated phytochrome signaling to specific basic helix- loop-helix transcription factors. Plant Cell, 2004,16:3033-3044.
doi: 10.1105/tpc.104.025643 pmid: 15486100 |
| [9] |
Ni M, Tepperman J M, Quail P H. Binding of phytochrome B to its nuclear signalling partner PIF3 is reversibly induced by light. Nature, 1999,400:781-784.
doi: 10.1038/23500 pmid: 10466729 |
| [10] |
Huq E, Al-Sady B, Hudson M, Kim C, Apel K, Quail P H. PHYTOCHROME-INTERACTING FACTOR 1 is a critical bHLH regulator of chlorophyll biosynthesis. Science, 2004,305:1937-1941.
pmid: 15448264 |
| [11] |
Huq E, Quail P H. PIF4, a phytochrome-interacting bHLH factor, functions as a negative regulator of phytochrome B signaling in Arabidopsis. EMBO J, 2002,21:2441-2450.
doi: 10.1093/emboj/21.10.2441 pmid: 12006496 |
| [12] |
Leivar P, Monte E, Al-Sady B, Carle C, Storer A, Alonso J M, Ecker J R, Quail P H. The Arabidopsis phytochrome-interacting factor PIF7, together with PIF3 and PIF4, regulates responses to prolonged red light by modulating phyB levels. Plant Cell, 2008,20:337-352.
pmid: 18252845 |
| [13] |
Quint M, Delker C, Franklin K A, Wigge P A, Halliday K J, van Zanten M. Molecular and genetic control of plant thermomorphogenesis. Nat Plants, 2016,2:15190.
doi: 10.1038/nplants.2015.190 pmid: 27250752 |
| [14] |
Casal J J, Balasubramanian S. Thermomorphogenesis. Annu Rev Plant Biol, 2019,70:321-346.
pmid: 30786235 |
| [15] |
Legris M, Klose C, Burgie E S, Rojas C C R, Neme M, Hiltbrunner A, Wigge P A, Schäfer E, Vierstra R D, Casal J J. Phytochrome B integrates light and temperature signals in Arabidopsis. Science, 2016,354:897-900.
pmid: 27789798 |
| [16] |
Jung J H, Domijan M, Klose C, Biswas S, Ezer D, Gao M, Khattak A K, Box M S, Charoensawan V, Cortijo S, Kumar M, Grant A, Locke J C W, Schäfer E, Jaeger K E, Wigge P A. Phytochromes function as thermosensors in Arabidopsis. Science, 2016,354:886-889.
pmid: 27789797 |
| [17] |
Box M S, Huang B E, Domijan M, Jaeger K E, Khattak A K, Yoo S J, Sedivy E L, Jones D M, Hearn T J, Webb A A R, Grant A, Locke J C W, Wigge P A. ELF3 controls thermoresponsive growth in Arabidopsis. Curr Biol, 2015,25:194-199.
doi: 10.1016/j.cub.2014.10.076 pmid: 25557663 |
| [18] |
Tasset C, Singh Yadav A, Sureshkumar S, Singh R, van der Woude L, Nekrasov M, Tremethick D, van Zanten M, Balasubramanian S. POWERDRESS-mediated histone deacetylation is essential for thermomorphogenesis in Arabidopsis thaliana. PLoS Genet, 2018,14:e1007280.
pmid: 29547672 |
| [19] |
Lee C M, Thomashow M F. Photoperiodic regulation of the C-repeat binding factor (CBF) cold acclimation pathway and freezing tolerance in Arabidopsis thaliana. Proc Natl Acad Sci USA, 2012,109:15054-15059.
pmid: 22927419 |
| [20] |
Liu Z J, Zhang Y Q, Wang J F, Li P, Zhao C Z, Chen Y D, Bi Y R. Phytochrome-interacting factors PIF4 and PIF5 negatively regulate anthocyanin biosynthesis under red light in Arabidopsis seedlings. Plant Sci, 2015,238:64-72.
pmid: 26259175 |
| [21] |
Gangappa S N, Berriri S, Kumar S V. PIF4 coordinates thermosensory growth and immunity in arabidopsis. Curr Biol, 2017,27:243-249.
doi: 10.1016/j.cub.2016.11.012 pmid: 28041792 |
| [22] |
Sun J Q, Qi L L, Li Y N, Chu J F, Li C Y. PIF4-mediated activation of YUCCA8 expression integrates temperature into the auxin pathway in regulating Arabidopsis hypocotyl growth. PLoS Genet, 2012,8:e1002594.
pmid: 22479194 |
| [23] |
Franklin K A, Lee S H, Patel D, Kumar S V, Spartz A K, Gu C, Ye S Q, Yu P, Breen G, Cohen J D, Wigge P A, Gray W M. Phytochrome-interacting factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc Natl Acad Sci USA, 2011,108:20231-20235.
pmid: 22123947 |
| [24] |
Hornitschek P, Kohnen M V, Lorrain S, Rougemont J, Ljung K, Lopez-Vidriero I, Franco-Zorrilla J M, Solano R, Trevisan M, Pradervand S, Xenarios I, Fankhauser C. Phytochrome interacting factors 4 and 5 control seedling growth in changing light conditions by directly controlling auxin signaling. Plant J, 2012,71:699-711.
pmid: 22536829 |
| [25] | Oh E, Zhu J Y, Bai M Y, Arenhart R A, Sun Y, Wang Z Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. eLife, 2014,3:e03031. |
| [26] |
Boure N, Kumar S V, Arnaud N. The BAP Module: a multisignal integrator orchestrating growth. Trends Plant Sci, 2019,24:602-610.
doi: 10.1016/j.tplants.2019.04.002 pmid: 31076166 |
| [27] |
Min L, Li Y Y, Hu Q, Zhu L F, Gao W H, Wu Y L, Ding Y H, Liu S M, Yang X Y, Zhang X L. Sugar and auxin signaling pathways respond to high-temperature stress during anther development as revealed by transcript profiling analysis in cotton. Plant Physiol, 2014,164:1293-1308.
pmid: 24481135 |
| [28] |
Min L, Zhu L F, Tu L L, Deng F L, Yuan D J, Zhang X L. Cotton GhCKI disrupts normal male reproduction by delaying tapetum programmed cell death via inactivating starch synthase. Plant J, 2013,75:823-835.
doi: 10.1111/tpj.12245 pmid: 23662698 |
| [29] | Min L, Hu Q, Li Y Y, Xu J, Ma Y Z, Zhu L F, Yang X Y, Zhang X L. LEAFY COTYLEDON1-CASEIN KINASE I-TCP15-PHYT OCHROME INTERACTING FACTOR4 network regulates somatic embryogenesis by regulating auxin homeostasis. Plant Physiol, 2015,169:2805-2821. |
| [30] | 胡海燕. 棉花茉莉酸信号负调控因子GhJAZ2的克隆及功能分析. 华中农业大学博士学位论文, 湖北武汉, 2016. |
| Hu H Y. The Cloning and Functional Characterization of a Cotton Negative Regulater GhJAZ2 in Jasmonic Acid Pathway. PhD Dissertation of Huazhong Agricultural University, Wuhan, Hubei, China, 2016 (in Chinese with English abstract). | |
| [31] | Jin S X, Zhang X L, Nie Y C, Guo X P, Liang S G, Zhu H G. Identification of a novel elite genotype for in vitro culture and genetic transformation of cotton. Biol Plant, 2006,50:519-524. |
| [32] |
Wu Y L, Min L, Wu Z C, Yang L, Zhu L F, Yang X Y, Yuan D J, Guo X P, Zhang X L. Defective pollen wall contributes to male sterility in the male sterile line 1355A of cotton. Sci Rep, 2015,5:9608.
doi: 10.1038/srep09608 pmid: 26043720 |
| [33] |
Yao X, Tian L, Yang J, Zhao Y N, Zhu Y X, Dai X, Zhao Y, Yang Z N. Auxin production in diploid microsporocytes is necessary and sufficient for early stages of pollen development. PLoS Genet, 2018,14:e1007397.
doi: 10.1371/journal.pgen.1007397 pmid: 29813066 |
| [34] |
Feng X L, Ni W M, Elge S, Mueller-Roeber B, Xu Z H, Xue H W. Auxin flow in anther filaments is critical for pollen grain development through regulating pollen mitosis. Plant Mol Biol, 2006,61:215-226.
doi: 10.1007/s11103-006-0005-z pmid: 16786302 |
| [35] |
Sun J Q, Qi L L, Li Y N, Zhai Q Z, Li C Y. PIF4 and PIF5 transcription factors link blue light and auxin to regulate the phototropic response in Arabidopsis. Plant Cell, 2013,25:2102-2114.
doi: 10.1105/tpc.113.112417 pmid: 23757399 |
| [36] |
Cheng Y F, Dai X H, Zhao Y D. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev, 2006,20:1790-1799.
pmid: 16818609 |
| [37] |
Tadashi S, Takeshi O, Shinya M, Mari T, Yuta T, Nahoko H, Yutaka M, Hideyuki T, Masao W, Atsushi H. Auxins reverse plant male sterility caused by high temperatures. Proc Natl Acad Sci USA, 2010,107:8569-8574.
doi: 10.1073/pnas.1000869107 pmid: 20421476 |
| [38] |
Cecchetti V, Altamura M M, Brunetti P, Petrocelli V, Falasca G, Ljung K, Costantino P, Cardarelli M. Auxin controls Arabidopsis anther dehiscence by regulating endothecium lignification and jasmonic acid biosynthesis. Plant J, 2013,74:411-422.
pmid: 23410518 |
| [39] |
Cecchetti V, Altamura M M, Falasca G, Costantino P, Cardarelli M. Auxin regulates Arabidopsis anther dehiscence, pollen maturation, and filament elongation. Plant Cell, 2008,20:1760-1774.
doi: 10.1105/tpc.107.057570 pmid: 18628351 |
| [40] |
Goldberg R B, Beals T P, Sanders P M. Anther development: basic principles and practical applications. Plant Cell, 1993,5:1217-1229.
doi: 10.1105/tpc.5.10.1217 pmid: 8281038 |
| [41] |
Yang C Y, Xu Z Y, Song J, Conner K, Vizcay Barrena G, Wilson Z A. Arabidopsis MYB26/MALE STERILE35 regulates secondary thickening in the endothecium and is essential for anther dehiscence. Plant Cell, 2007,19:534-548.
doi: 10.1105/tpc.106.046391 pmid: 17329564 |
| [42] |
Stepanova A N, Robertson-Hoyt J, Yun J, Benavente L M, Xie D Y, Dolezal K, Schlereth A, Jurgens G, Alonso J M. TAA1-mediated auxin biosynthesis is essential for hormone crosstalk and plant development. Cell, 2008,133:177-191.
doi: 10.1016/j.cell.2008.01.047 pmid: 18394997 |
| [43] |
Muller T M, Bottcher C, Morbitzer R, Gotz C C, Lehmann J, Lahaye T, Glawischnig E. TRANSCRIPTION ACTIVATOR- LIKE EFFECTOR NUCLEASE-mediated generation and metabolic analysis of camalexin-deficient cyp71a12 cyp71a13 double knockout lines. Plant Physiol, 2015,168:849-858.
doi: 10.1104/pp.15.00481 pmid: 25953104 |
| [44] |
Nagpal P, Ellis C M, Weber H, Ploense S E, Barkawi L S, Guilfoyle T J, Hagen G, Alonso J M, Cohen J D, Farmer E E, Ecker J R, Reed J W. Auxin response factors ARF6 and ARF8 promote jasmonic acid production and flower maturation. Development, 2005,132:4107-4118.
doi: 10.1242/dev.01955 pmid: 16107481 |
| [45] |
Wu M F, Tian Q, Reed J W. Arabidopsis microRNA167 controls patterns of ARF6 and ARF8 expression, and regulates both female and male reproduction. Development, 2006,133:4211-4218.
pmid: 17021043 |
| [1] | ZHOU Jing-Yuan, KONG Xiang-Qiang, ZHANG Yan-Jun, LI Xue-Yuan, ZHANG Dong-Mei, DONG He-Zhong. Mechanism and technology of stand establishment improvements through regulating the apical hook formation and hypocotyl growth during seed germination and emergence in cotton [J]. Acta Agronomica Sinica, 2022, 48(5): 1051-1058. |
| [2] | SUN Si-Min, HAN Bei, CHEN Lin, SUN Wei-Nan, ZHANG Xian-Long, YANG Xi-Yan. Root system architecture analysis and genome-wide association study of root system architecture related traits in cotton [J]. Acta Agronomica Sinica, 2022, 48(5): 1081-1090. |
| [3] | YAN Xiao-Yu, GUO Wen-Jun, QIN Du-Lin, WANG Shuang-Lei, NIE Jun-Jun, ZHAO Na, QI Jie, SONG Xian-Liang, MAO Li-Li, SUN Xue-Zhen. Effects of cotton stubble return and subsoiling on dry matter accumulation, nutrient uptake, and yield of cotton in coastal saline-alkali soil [J]. Acta Agronomica Sinica, 2022, 48(5): 1235-1247. |
| [4] | ZHENG Shu-Feng, LIU Xiao-Ling, WANG Wei, XU Dao-Qing, KAN Hua-Chun, CHEN Min, LI Shu-Ying. On the green and light-simplified and mechanized cultivation of cotton in a cotton-based double cropping system [J]. Acta Agronomica Sinica, 2022, 48(3): 541-552. |
| [5] | ZHANG Yan-Bo, WANG Yuan, FENG Gan-Yu, DUAN Hui-Rong, LIU Hai-Ying. QTLs analysis of oil and three main fatty acid contents in cottonseeds [J]. Acta Agronomica Sinica, 2022, 48(2): 380-395. |
| [6] | ZHANG Te, WANG Mi-Feng, ZHAO Qiang. Effects of DPC and nitrogen fertilizer through drip irrigation on growth and yield in cotton [J]. Acta Agronomica Sinica, 2022, 48(2): 396-409. |
| [7] | ER Chen, LIN Tao, XIA Wen, ZHANG Hao, XU Gao-Yu, TANG Qiu-Xiang. Coupling effects of irrigation and nitrogen levels on yield, water distribution and nitrate nitrogen residue of machine-harvested cotton [J]. Acta Agronomica Sinica, 2022, 48(2): 497-510. |
| [8] | JIAN Hong-Ju, SHANG Li-Na, JIN Zhong-Hui, DING Yi, LI Yan, WANG Ji-Chun, HU Bai-Geng, Vadim Khassanov, LYU Dian-Qiu. Genome-wide identification and characterization of PIF genes and their response to high temperature stress in potato [J]. Acta Agronomica Sinica, 2022, 48(1): 86-98. |
| [9] | ZHAO Wen-Qing, XU Wen-Zheng, YANG Liu-Yan, LIU Yu, ZHOU Zhi-Guo, WANG You-Hua. Different response of cotton leaves to heat stress is closely related to the night starch degradation [J]. Acta Agronomica Sinica, 2021, 47(9): 1680-1689. |
| [10] | YUE Dan-Dan, HAN Bei, Abid Ullah, ZHANG Xian-Long, YANG Xi-Yan. Fungi diversity analysis of rhizosphere under drought conditions in cotton [J]. Acta Agronomica Sinica, 2021, 47(9): 1806-1815. |
| [11] | ZENG Zi-Jun, ZENG Yu, YAN Lei, CHENG Jin, JIANG Cun-Cang. Effects of boron deficiency/toxicity on the growth and proline metabolism of cotton seedlings [J]. Acta Agronomica Sinica, 2021, 47(8): 1616-1623. |
| [12] | GAO Lu, XU Wen-Liang. GhP4H2 encoding a prolyl-4-hydroxylase is involved in regulating cotton fiber development [J]. Acta Agronomica Sinica, 2021, 47(7): 1239-1247. |
| [13] | LI Wen-Lan, LI Wen-Cai, SUN Qi, YU Yan-Li, ZHAO Meng, LU Shou-Ping, LI Yan-Jiao, MENG Zhao-Dong. A study of expression pattern of auxin response factor family genes in maize (Zea mays L.) [J]. Acta Agronomica Sinica, 2021, 47(6): 1138-1148. |
| [14] | MA Huan-Huan, FANG Qi-Di, DING Yuan-Hao, CHI Hua-Bin, ZHANG Xian-Long, MIN Ling. GhMADS7 positively regulates petal development in cotton [J]. Acta Agronomica Sinica, 2021, 47(5): 814-826. |
| [15] | XU Nai-Yin, ZHAO Su-Qin, ZHANG Fang, FU Xiao-Qiong, YANG Xiao-Ni, QIAO Yin-Tao, SUN Shi-Xian. Retrospective evaluation of cotton varieties nationally registered for the Northwest Inland cotton growing regions based on GYT biplot analysis [J]. Acta Agronomica Sinica, 2021, 47(4): 660-671. |
|
||