Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2019, Vol. 45 ›› Issue (7): 1017-1028.doi: 10.3724/SP.J.1006.2019.84142
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
TIAN Wen-Gang1,ZHU Xue-Feng1,SONG Wen1,CHENG Wen-Han2,XUE Fei1,ZHU Hua-Guo1,*( )
)
| [1] |
Alcázar R, Altabella T, Marco F, Bortolotti C, Reymond M, Koncz C, Carrasco P, Tiburcio A F . Polyamines: molecules with regulatory functions in plant abiotic stress tolerance. Planta, 2010,231:1237-1249.
doi: 10.1007/s00425-010-1130-0 |
| [2] |
Kusano T, Berberich T C, Takahashi Y . Polyamines: essential factors for growth and survival. Planta, 2008,228:367-381.
doi: 10.1007/s00425-008-0772-7 |
| [3] |
Slocum R D, Kaur-Sawhney R, Galston A W . The physiology and biochemistry of polyamines in plants. Arch Biochem Biophys, 1984,235:283-303.
doi: 10.1016/0003-9861(84)90201-7 |
| [4] | Soo W, Soo K, Woo K, Ky P . Constitutive S-adenosylmethionine decarboxylase gene expression increases drought tolerance through inhibition of reactive oxygen species accumulation in Arabidopsis. Planta, 2014,239:979-988. |
| [5] | Torrigiani P, Scaramagli S, Ziosi V, Mayer M, Biondi S . Expression of an antisense Datura stramonium S-adenosylmethionine decarboxylase cDNA in tobacco: changes in enzyme activity, putrescine-spermidine ratio, rhizogenic potential, and response to methyl jasmonate. J Plant Physiol, 2005,162:559-571. |
| [6] |
Sinha R, Rajam M V . RNAi silencing of three homologues of S-adenosylmethionine decarboxylase gene in tapetal tissue of tomato results in male sterility. Plant Mol Biol, 2013,82:169-180.
doi: 10.1007/s11103-013-0051-2 |
| [7] |
路玉兰, 孙艳香, 冯雪, 赵学良 . 百脉根S-腺苷甲硫氨酸脱羧酶基因克隆与表达分析. 华北农学报, 2013,28(2):78-85.
doi: 10.3969/j.issn.1000-7091.2013.02.015 |
|
Lu Y L, Sun Y X, Feng X, Zhao X L . Cloning and expression analysis of S-adenosylmethionine decarboxylase gene from Lotus corniculatus L. Acta Agric Boreali-Sin, 2013,28(2):78-85 (in Chinese with English abstract).
doi: 10.3969/j.issn.1000-7091.2013.02.015 |
|
| [8] | Peng X J, Zhang L X, Zhang L X, Liu Z J, Cheng L Q, Yang Y, Shen S H, Chen S Y, Liu G S . The transcriptional factor LcDREB2 cooperates with LcSAMDC2 to contribute to salt tolerance in Leymus chinensis. Plant Cell Tissue Organ Culture, 2013,113:245-256. |
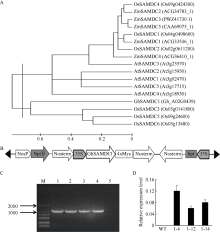
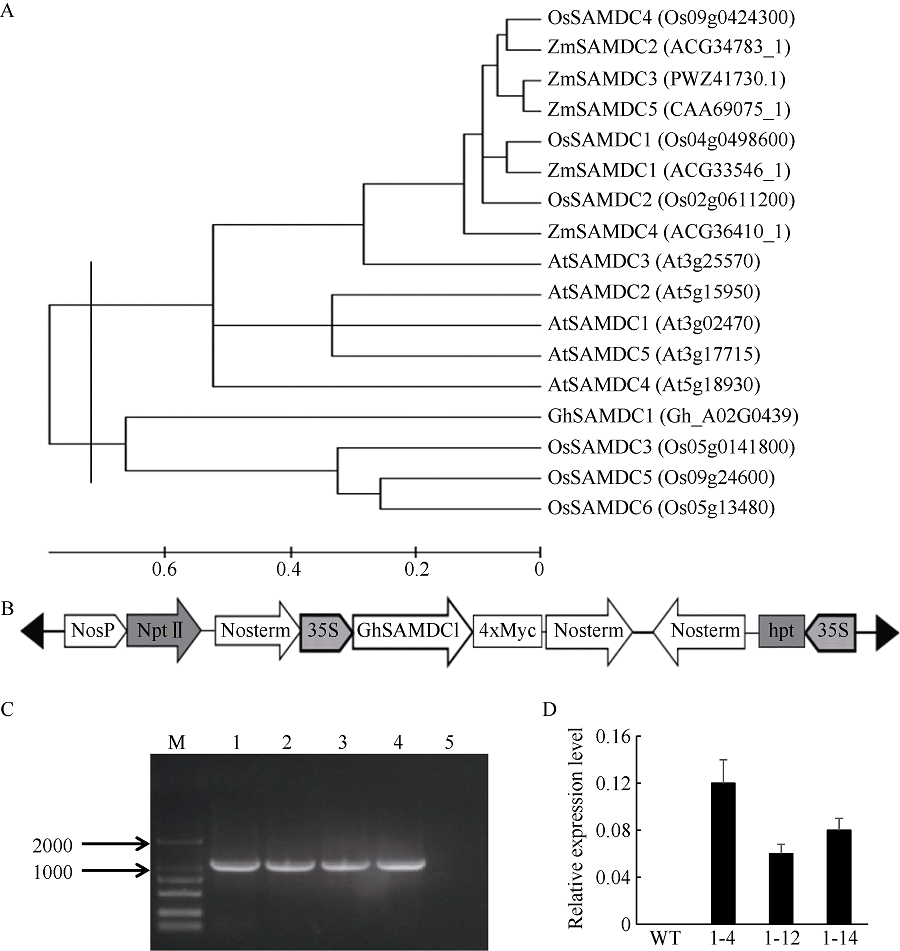
| [9] | 王凡龙, 朱华国, 程文翰, 刘永昌, 成新琪, 孙杰 . 棉花S-腺苷蛋氨酸脱羧酶基因(GhSAMDC2/3/4)的克隆及其诱导表达分析. 棉花学报, 2015,27:176-183. |
| Wang F L, Zhu H G, Cheng W H, Liu Y C, Cheng X Q, Sun J . Cloning and induced expression analysis ofGhSAMDC2/3/4 in cotton(Gossypium hirsutum L.). Cotton Sci, 2015,27:176-183 (in Chinese with English abstract). | |
| [10] |
张梅, 王然, 马春晖, 段艳欣, 李鼎立 . 杜梨S-腺苷甲硫氨酸脱羧酶基因的克隆与生物信息学分析. 华北农学报, 2013,28(1):82-87.
doi: 10.3969/j.issn.1000-7091.2013.01.016 |
|
Zhang M, Wang R, Ma C H, Duan Y X, Li D L . Cloning and bioinformatics analysis of S-adenosylmethionine decarboxylase gene in Pyrus betulaefolia Bge. Acta Agric Boreali-Sin, 2013,28(1):82-87 (in Chinese with English abstract).
doi: 10.3969/j.issn.1000-7091.2013.01.016 |
|
| [11] | 文乐, 黄诚梅, 邓智年, 曹辉庆, 魏源文, 李楠, 吴凯朝 . 甘蔗S-腺苷蛋氨酸脱羧酶基因Sc-SAMDC3的克隆和表达分析. 南方农业学报, 2015,46:1931-1936. |
| Wen Y, Huang C M, Deng Z N, Cao H Q, Li N, Wu K C . Molecular cloning of sugarcane S-adenosylmethionine decarboxylase gene (Sc-SAMDC3) and its expression analysis. J Southern Agric, 2015,46:1931-1936 (in Chinese with English abstract). | |
| [12] |
王小利, 刘晓霞, 王舒颖, 杨义成, 吴佳海 . 高羊茅腺苷甲硫氨酸脱羧酶基因FaSAMDC的克隆与差异表达分析. 草业学报, 2011,20(4):169-179.
doi: 10.11686/cyxb20110421 |
|
Wang X L, Liu X X, Wang S Y, Yang Y C, Wu J H . Cloning and differential expression analysis of S-adenosylmethionine decarboxylase gene FaSAMDC in tall fescue. Acta Pratac Sin, 2011,20(4):169-179 (in Chinese with English abstract).
doi: 10.11686/cyxb20110421 |
|
| [13] | Liu Z J, Liu P P, Qi D M, Peng X J, Liu G S . Enhancement of cold and salt tolerance of Arabidopsis by transgenic expression of the S-adenosylmethionine decarboxylase gene from Leymus chinensis. J Plant Physiol, 2017,211:90-99. |
| [14] | Cheng L, Zou Y J, Ding S L, Zhang J J, Yu X L, Cao J S, Lu G . Polyamine accumulation in transgenictomato enhances the tolerance to high temperature stress. Chin Bull Bot, 2009,51:489-499. |
| [15] |
Chen M, Chen J J, Fang J Y, Guo Z F, Lu S Y . Down-regulation of S-adenosylmethionine decarboxylase genes results in reduced plant length, pollen viability, and abiotic stress tolerance. Plant Cell Tissue Organ Culture, 2014,116:311-322.
doi: 10.1007/s11240-013-0405-0 |
| [16] | Elsayed A I, Rafudeen M S, El-hamahmy M A M, Odero D C, Sazzad H M . Enhancing antioxidant systems by exogenous spermine and spermidine in wheat (Triticum aestivum) seedlings exposed to salt stress. Funct Plant Biol, 2018,45:745, doi: 10.1071/FP17127. |
| [17] |
Wu J Q, Shu S, Li C C, Sun J, Guo S R . Spermidine-mediated hydrogen peroxide signaling enhances the antioxidant capacity of salt-stressed cucumber roots. Plant Physiol Biochem, 2018,128:152-162.
doi: 10.1016/j.plaphy.2018.05.002 |
| [18] | Zhou H, Guo S R, An Y H, Shan X, Wang Y, Shu S, Sun J . Exogenous spermidine delays chlorophyll metabolism in cucumber leaves (Cucumis sativus L.) under high temperature stress. Acta Physiol Planta, 2016,38:224. |
| [19] |
Liu J, Yu B J, Liu Y L . Effects of spermidine and spermine levels on salt tolerance associated with tonoplast H +-ATPase and H +-PPase activities in barley roots . Plant Growth Regul, 2006,49:119-126.
doi: 10.1007/s10725-006-9001-1 |
| [20] | Duan J J, Guo S R, Fan H F, Wang S P, Kang Y Y . Effects of salt stress on proline and polyamine metabolisms in the roots of cucumber seedlings. Acta Bot Boreali-Occident Sin, 2006,26:2486-2492. |
| [21] |
Halliwell B . Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol, 2006,141:312-322.
doi: 10.1104/pp.106.077073 |
| [22] | Ma C Q, Wang Y G, Gu D, Nan J D, Chen S X, Li H Y . Overexpression of S-adenosyl-l-methionine synthetase 2 from sugar beet M14 increased Arabidopsis tolerance to salt and oxidative stress. Int J Mol Sci, 2017,18:e847. |
| [23] |
Li J M, Hu L P, Zhang L, Pan X B, Hu X H . Exogenous spermidine is enhancing tomato tolerance to salinity-alkalinity stress by regulating chloroplast antioxidant system and chlorophyll metabolism. BMC Plant Biol, 2015,15:303, doi: 10.1186/s12870- 015-0699-7.
doi: 10.1186/s12870-015-0699-7 |
| [24] |
Asada K . THE WATER-WATER CYCLE IN CHLOROPLASTS: Scavenging of active oxygens and dissipation of excess photons. Annu Rev Plant Physiol Plant Mol Biol, 1999,50:601-639.
doi: 10.1146/annurev.arplant.50.1.601 |
| [25] |
Williamson G B, Richardson D . Bioassays for allelopathy: Measuring treatment responses with independent controls. J Chem Ecol, 1988,14:181-187.
doi: 10.1007/BF01022540 |
| [26] |
Zhao F Y, Guo S L, Zhang H, Zhao Y X . Expression of yeast SOD2, in transgenic rice results in increased salt tolerance. Plant Sci, 2006,170:216-224.
doi: 10.1016/j.plantsci.2005.08.017 |
| [27] | Puyang X H, An M Y, Han L B, Zhang X Z . Protective effect of spermidine on salt stress induced oxidative damage in two Kentucky bluegrass (Poa pratensis L.) cultivars. Ecotoxicol Environ Saf, 2015,117:96-106. |
| [28] | Li S, Han J, Qiang Z . The effect of exogenous spermidine concentration on polyamine metabolism and salt tolerance in Zoysiagrass (Zoysia japonica Steud) subjected to short-term salinity stress. Front Plant Sci, 2016,7:1221, doi: 10.3389/fpls. 2016.01221. |
| [29] |
Radhakrishnan R . Ameliorative effects of spermine against osmotic stress through antioxidants and abscisic acid changes in soybean pods and seeds. Acta Physiol Planta, 2013,35:263-269.
doi: 10.1007/s11738-012-1072-1 |
| [30] |
Zrig A, Tounelti T, Vadel A M, Mohamed H B, Valero D, Serrano M, Chtara C, Khemira H . Possible involvement of polyphenols and polyamines in salt tolerance of almond rootstocks. Plant Physiol Biochem, 2011,49:1313-1322.
doi: 10.1016/j.plaphy.2011.08.009 |
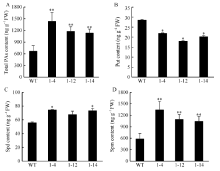
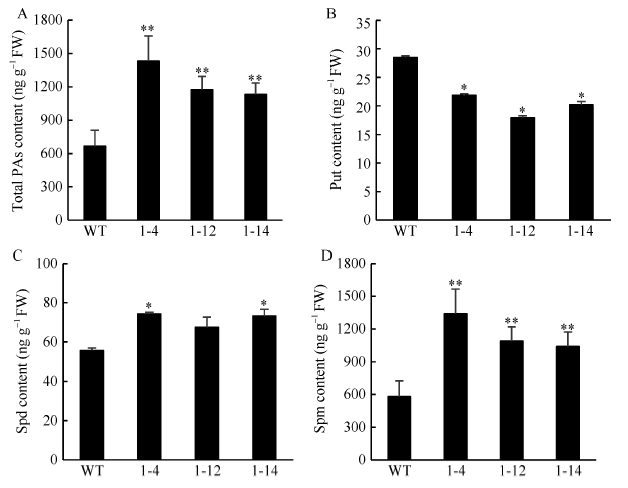
| [31] | 程文翰, 朱华国, 李鹏飞, 王凡龙, 朱守鸿, 赵兰杰, 郭丽雪, 孙杰 . 棉花多胺HPLC的测定方法优化及其在体细胞胚胎发生过程中的变化规律. 棉花学报, 2014,26:138-144. |
| Cheng W H, Zhu H G, Li P F, Wang F L, Zhu S H, Zhao L J, Guo L X, Sun J . Method optimization of polyamine content by high-performance liquid chromatography and its changes in the process of somatic embryogenesis in cotton. Cotton Sci, 2014,26:138-144 (in Chinese with English abstract). | |
| [32] | Cheng W H, Wang F H, Cheng X Q, Zhu Q H, Sun Y Q, Zhu H G, Sun J . Polyamine and its metabolite H2O2 play a key role in the conversion of embryogenic callus into somatic embryos in upland cotton (Gossypium hirsutum L.). Front Plant Sci, 2015,6:1063, doi: 10.3389/fpls.2015.01063. |
| [33] | Li C, Zhang Y N, Zhang K, Guo D L, Cui B M, Wang X Y, Huang X Z . Promoting flowering, lateral shoot outgrowth, leaf development, and flower abscission in tobacco plants overexpressing cotton FLOWERING LOCUS T (FT)-like gene GhFT1. Front Plant Sci, 2015,6:454, doi: 10.3389/fpls.2015.00454. |
| [34] |
Crumbliss A L, Perine S C, Stonehuerner J, Tubergen K R, Zhao J, Henkens R W, Q’Daly J P . Colloidal gold as a biocompatible immobilization matrix suitable for the fabrication of enzyme electrodes by electrodeposition. Biotechnol Bioeng, 1992,40:483-490.
doi: 10.1002/(ISSN)1097-0290 |
| [35] | 朱珍 . 赤霉素调控采后番茄果实抗冷机制研究. 中国农业科学院硕士学位论文, 北京, 2016. |
| Zhu Z . The Mechanism of Gibberellins in Regulation of Chilling Tolerance of Postharvest Tomato Fruit. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2016 (in Chinese with English abstract). | |
| [36] |
Livak K J, Schmittgen T D . Analysis of relative gene expression data using real-time quantitative PCR and the 2 -ΔΔCT method . Methods, 2001,25:402-408.
doi: 10.1006/meth.2001.1262 |
| [37] |
Hao Y J, Zhang Z L, Kitashiba H, Honda C, Ubi B, Kita M, Moriguchi T . Molecular cloning and functional characterization of two apple S-adenosylmethionine decarboxylase genes and their different expression in fruit development, cell growth and stress responses. Gene, 2005,350:41-50.
doi: 10.1016/j.gene.2005.01.004 |
| [38] |
Roy M, Wu R . Overexpression of S-adenosylmethionine decarboxylase gene in rice increases polyamine level and enhances sodium chloride-stress tolerance. Plant Sci, 2002,163:987-992.
doi: 10.1016/S0168-9452(02)00272-8 |
| [39] | Waie B, Rajam M V . Effect of increased polyamine biosynthesis on stress responses in transgenic tobacco by introduction of human S-adenosylmethionine gene. Plant Sci, 2003,164:722-734. |
| [40] | Sanchez D H, Cuevas J C, Chiesa M A, Ruiz O A . Free spermidine and spermine content in Lotus glaber under long-term salt stress. Plant Sci, 2005,168:541-546. |
| [41] |
Zapata P J, Serrano M, Pretel M T, Amoros A, Botella M A . Polyamines and ethylene changes during germination of different plant species under salinity. Plant Sci, 2004,167:781-788.
doi: 10.1016/j.plantsci.2004.05.014 |
| [42] |
Liu H P, Dong B H, Zhang Y Y, Liu Z P, Liu Y L . Relationship between osmotic stress and the levels of free, conjugated and bound polyamines in leaves of wheat seedlings. Plant Sci, 2004,166:1261-1267.
doi: 10.1016/j.plantsci.2003.12.039 |
| [43] | Wi S J, Kim W T, Park K Y . Overexpression of camation S-adenosylmethionine decarboxylase gene generate a broad- spectrum tolerance to abiotic stresses in transgenenic tobacco plants. Plant Cell Rep, 2006,25:1111-1121. |
| [44] | Liu H P, Zhu Z X, Liu Y L . Response of bound polyamines in the thylakoid membrane of wheat seedling to osmotic stress. Seed, 2007,26:58-60. |
| [45] |
Ha H C, Sirisoma N S, Kuppusamy P, Zweier J L, Woster P M, Casero R A . The natural polyamine spermine functions directly as a free radical scavenger. Proc Natl Acad Sci USA, 1998,95:11140-11145.
doi: 10.1073/pnas.95.19.11140 |
| [46] |
Tiburcio A F, Besford R T, Capell T, Borrell A, Testillano P S, Risueno M C . Mechanisms of polyamine action during senescence responses induced by osmotic stress. J Exp Bot, 1994,45:1789-1800.
doi: 10.1093/jxb/45.12.1789 |
| [1] | YAN Jia-Qian, GU Yi-Biao, XUE Zhang-Yi, ZHOU Tian-Yang, GE Qian-Qian, ZHANG Hao, LIU Li-Jun, WANG Zhi-Qin, GU Jun-Fei, YANG Jian-Chang, ZHOU Zhen-Ling, XU Da-Yong. Different responses of rice cultivars to salt stress and the underlying mechanisms [J]. Acta Agronomica Sinica, 2022, 48(6): 1463-1475. |
| [2] | LEI Xin-Hui, WAN Chen-Xi, TAO Jin-Cai, LENG Jia-Jun, WU Yi-Xin, WANG Jia-Le, WANG Peng-Ke, YANG Qing-Hua, FENG Bai-Li, GAO Jin-Feng. Effects of soaking seeds with MT and EBR on germination and seedling growth in buckwheat under salt stress [J]. Acta Agronomica Sinica, 2022, 48(5): 1210-1221. |
| [3] | MENG Ying, XING Lei-Lei, CAO Xiao-Hong, GUO Guang-Yan, CHAI Jian-Fang, BEI Cai-Li. Cloning of Ta4CL1 and its function in promoting plant growth and lignin deposition in transgenic Arabidopsis plants [J]. Acta Agronomica Sinica, 2022, 48(1): 63-75. |
| [4] | DAI Liang-Xiang, XU Yang, ZHANG Guan-Chu, SHI Xiao-Long, QIN Fei-Fei, DING Hong, ZHANG Zhi-Meng. Response of rhizosphere bacterial community diversity to salt stress in peanut [J]. Acta Agronomica Sinica, 2021, 47(8): 1581-1592. |
| [5] | LI Zeng-Qiang, DING Xin-Chao, LU Hai, HU Ya-Li, YUE Jiao, HUANG Zhen, MO Liang-Yu, CHEN Li, CHEN Tao, CHEN Peng. Physiological characteristics and DNA methylation analysis under lead stress in kenaf (Hibiscus cannabinus L.) [J]. Acta Agronomica Sinica, 2021, 47(6): 1031-1042. |
| [6] | LIU Ya-Wen, ZHANG Hong-Yan, CAO Dan, LI Lan-Zhi. Prediction of drought and salt stress-related genes in rice based on multi-platform gene expression data [J]. Acta Agronomica Sinica, 2021, 47(12): 2423-2439. |
| [7] | Hui LI, De-Fang LI, Yong DENG, Gen PAN, An-Guo CHEN, Li-Ning ZHAO, Hui-Juan TANG. Cloning of the key enzyme gene HcTPPJ in trehalose biosynthesis of kenaf and its expression in response to abiotic stress in kenaf [J]. Acta Agronomica Sinica, 2020, 46(12): 1914-1922. |
| [8] | LI Run-Zhi, JIN Qing, LI Zhao-Hu, WANG Ye, PENG Zhen, DUAN Liu-Sheng. Salicylic acid improved salinity tolerance of Glycyrrhiza uralensis Fisch during seed germination and seedling growth stages [J]. Acta Agronomica Sinica, 2020, 46(11): 1810-1816. |
| [9] | CHEN Xiao-Jing,LIU Jing-Hui,YANG Yan-Ming,ZHAO Zhou,XU Zhong-Shan,HAI Xia,HAN Yu-Ting. Effects of salt stress on physiological indexes and differential proteomics of oat leaf [J]. Acta Agronomica Sinica, 2019, 45(9): 1431-1439. |
| [10] | LI Xu-Kai,LI Ren-Jian,ZHANG Bao-Jun. Identification of rice stress-related gene co-expression modules by WGCNA [J]. Acta Agronomica Sinica, 2019, 45(9): 1349-1364. |
| [11] | Ning HE,Xue-Yang WANG,Liang-Zi CAO,Da-Wei CAO,Yu LUO,Lian-Zi JIANG,Ying MENG,Chun-Xu LENG,Xiao-Dong TANG,Yi-Dan LI,Shu-Ming WAN,Huan LU,Xu-Zhen CHENG. Effects of photoperiods and temperatures on physiological characteristics and chlorophyll synthesis precursors of adzuki bean seedlings [J]. Acta Agronomica Sinica, 2019, 45(3): 460-468. |
| [12] | Hua-Ying MAO,Feng LIU,Wei-Hua SU,Ning HUANG,Hui LING,Xu ZHANG,Wen-Ju WANG,Cong-Na LI,Han-Chen TANG,Ya-Chun SU,You-Xiong QUE. A Sugarcane Phosphatidylinositol Transfer Protein Gene ScSEC14 Responds to Drought and Salt Stresses [J]. Acta Agronomica Sinica, 2018, 44(6): 824-835. |
| [13] | Xiang ZHAO,Zi-Yi ZHU,Xiao-Nan WANG,Shi-Chao MU,Xiao ZHANG. Functional Analysis of Hypocotyl Phototropism Modulated by RPT2-Interacting Protein RIP1 in Arabidopsis thaliana L. [J]. Acta Agronomica Sinica, 2018, 44(12): 1802-1808. |
| [14] | Guang-Long ZHU,Cheng-Yu SONG,Lin-Lin YU,Xu-Bing CHEN,Wen-Fang ZHI,Jia-Wei LIU,Xiu-Rong JIAO,Gui-Sheng ZHOU. Alleviation Effects of Exogenous Growth Regulators on Seed Germination of Sweet Sorghum under Salt Stress and Its Physiological Basis [J]. Acta Agronomica Sinica, 2018, 44(11): 1713-1724. |
| [15] | Li-Li WAN, Zhuan-Rong WANG, Qiang XIN, Fa-Ming DONG, Deng-Feng HONG, Guang-Sheng YANG. Enhanced Accumulation of BnA7HSP70 Molecular Chaperone Binding Protein Improves Tolerance to Drought Stress in Transgenic Brassica napus [J]. Acta Agronomica Sinica, 2018, 44(04): 483-492. |
|
||