欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (8): 2105-2121.doi: 10.3724/SP.J.1006.2023.24194
代书桃1( ), 朱灿灿1, 马小倩2, 秦娜1, 宋迎辉1, 魏昕1, 王春义1, 李君霞1,*()
), 朱灿灿1, 马小倩2, 秦娜1, 宋迎辉1, 魏昕1, 王春义1, 李君霞1,*()
DAI Shu-Tao1(), ZHU Can-Can1, MA Xiao-Qian2, QIN Na1, SONG Ying-Hui1, WEI Xin1, WANG Chun-Yi1, LI Jun-Xia1,*()
摘要:
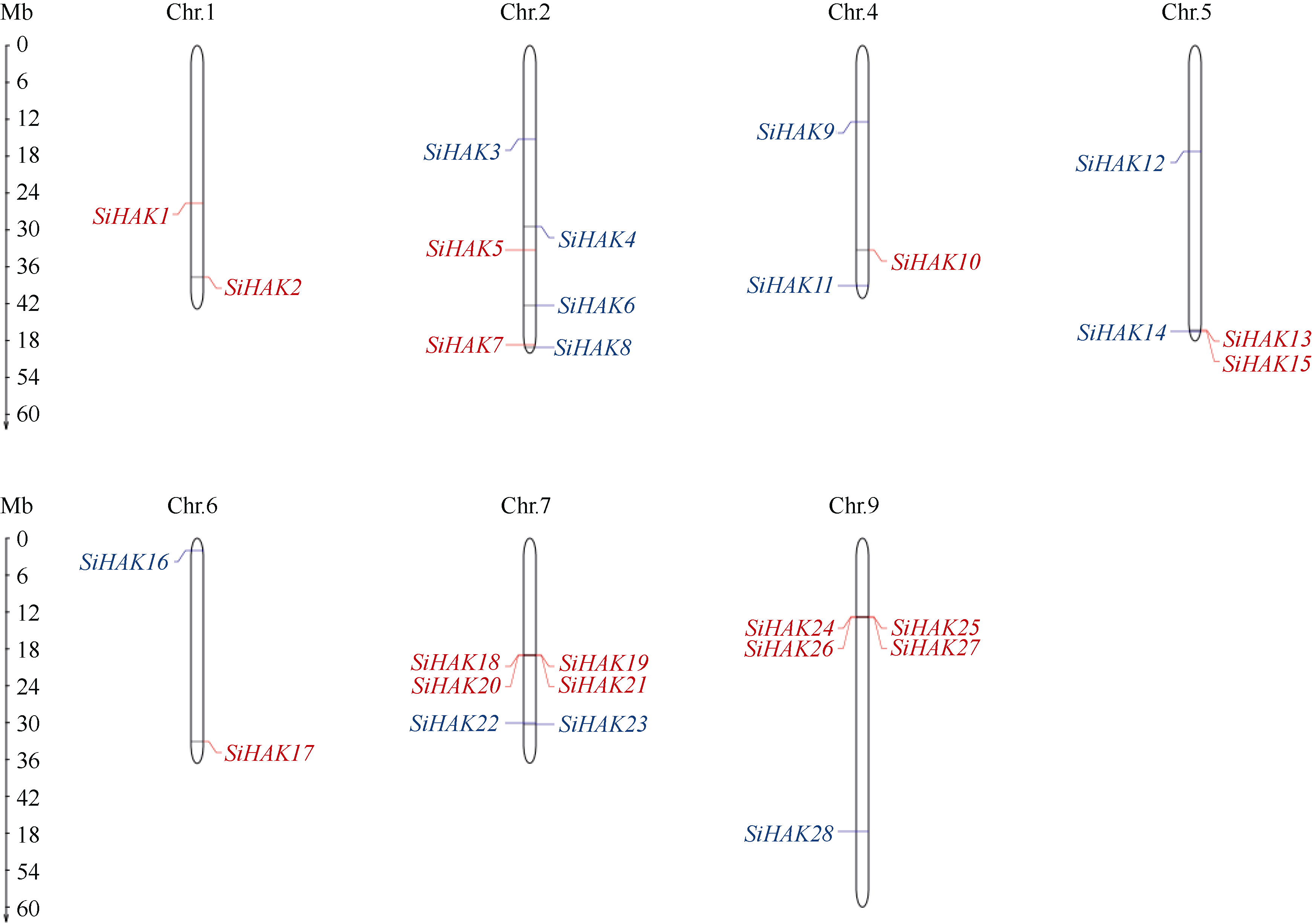
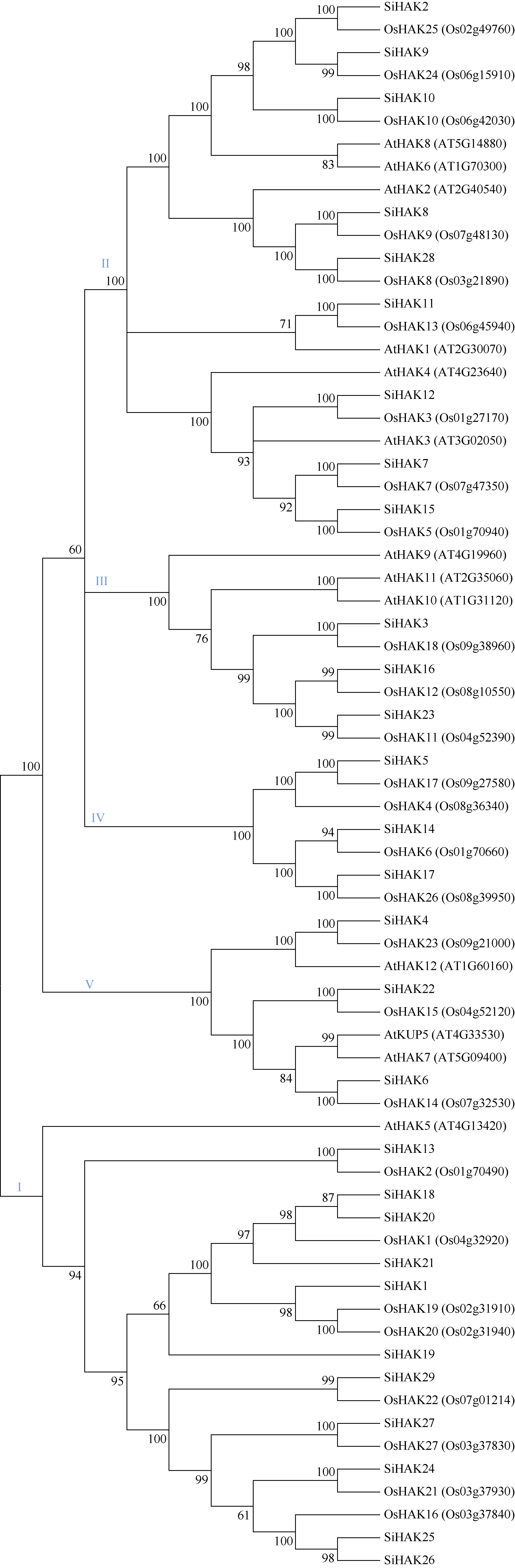
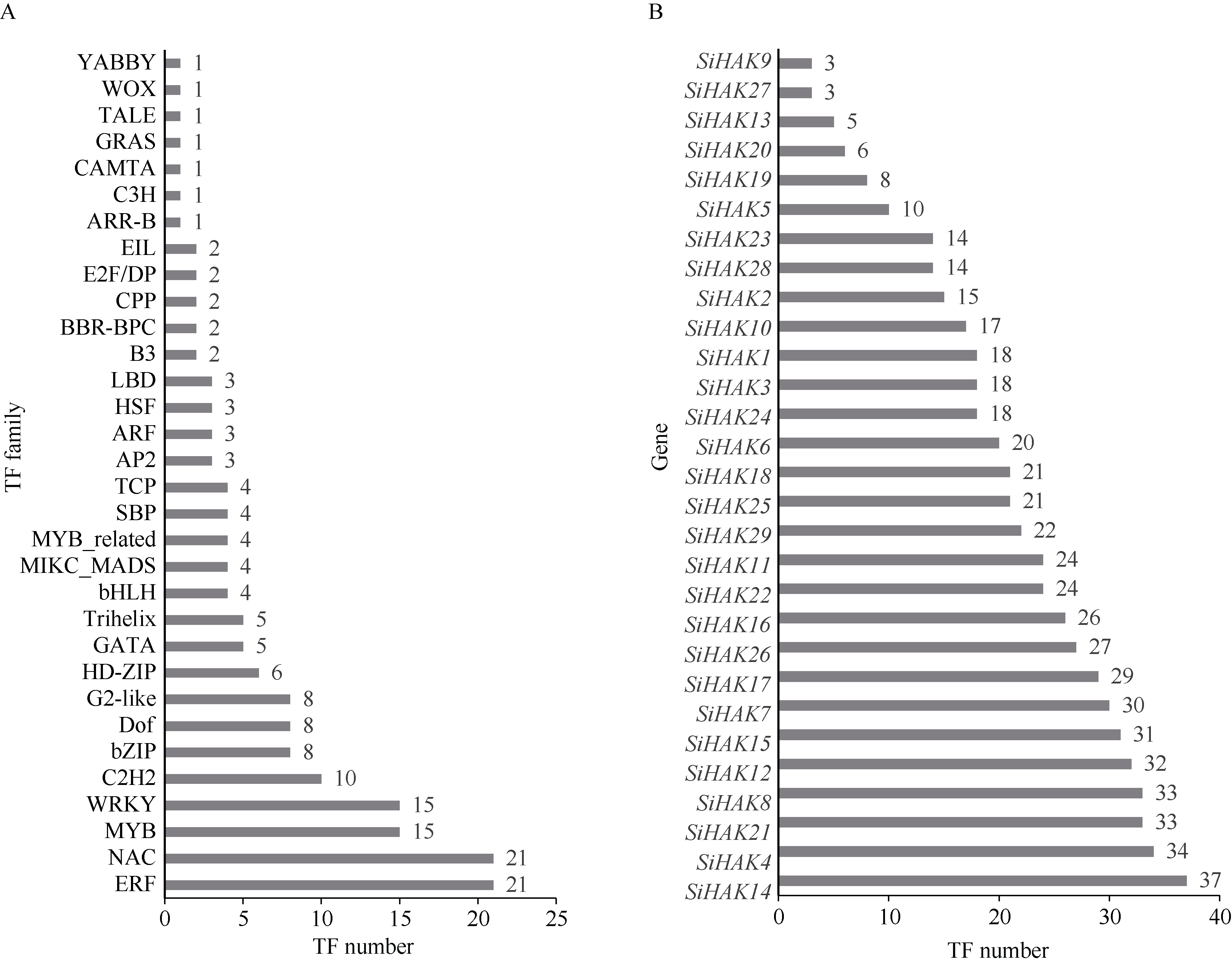
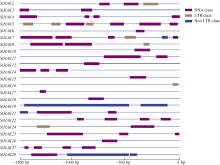
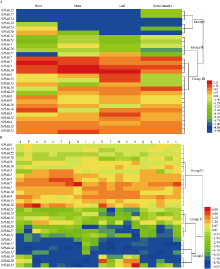
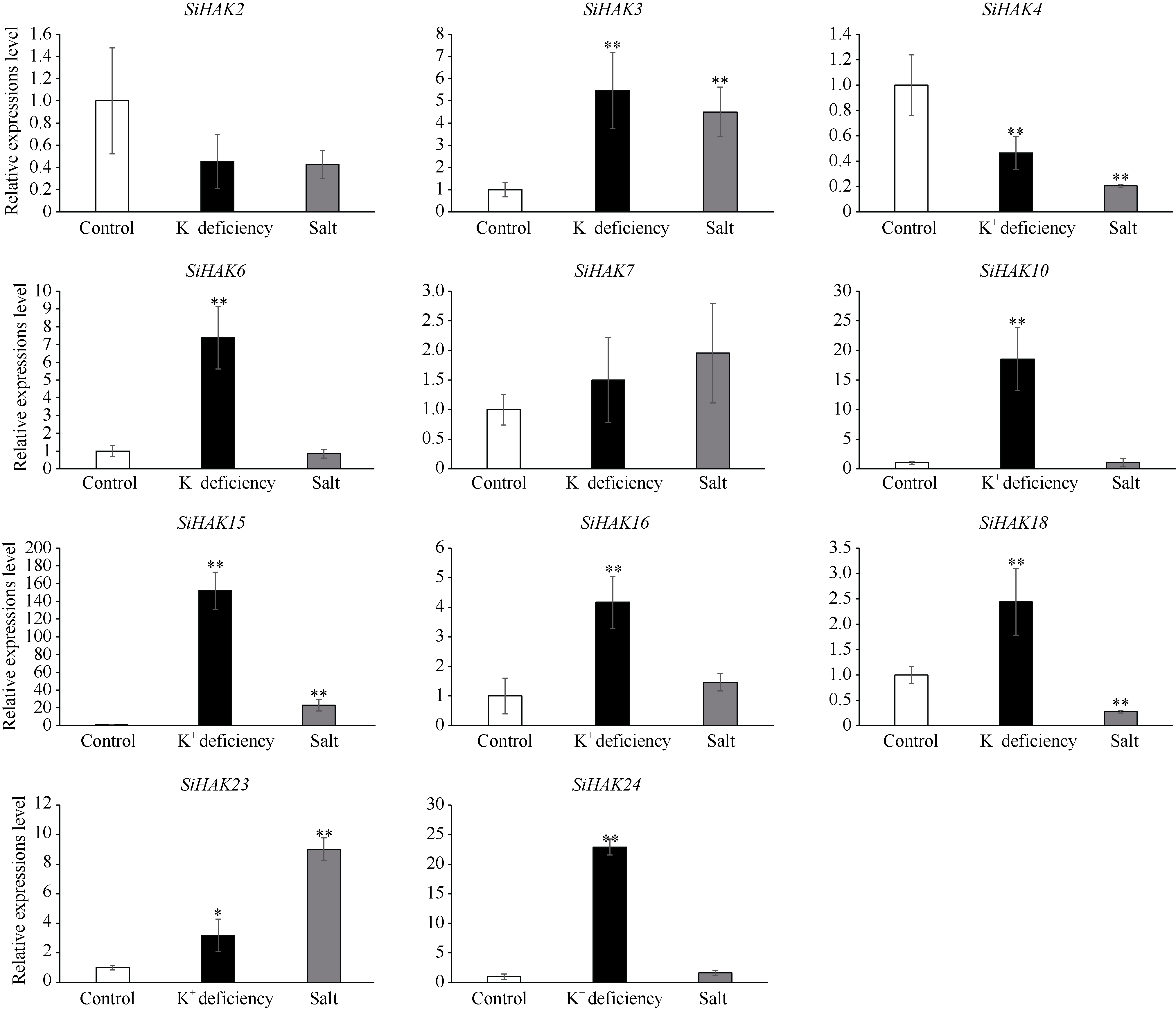
KT/HAK/KUP (HAK)家族是植物中最丰富的钾转运体家族, 对植物的生长和环境适应具有重要作用。谷子是抗逆耐瘠研究的模式植物, 然而, 谷子中HAK家族缺乏系统研究。本研究基于基因组序列信息, 鉴定出29个谷子HAK基因(SiHAKs), 并对该家族成员的基本特征、蛋白结构、染色体定位、基因复制、表达模式和逆境响应等方面进行了系统分析。结果显示, (1) SiHAKs分为5个进化簇(Cluster I~Cluster V), 成员数量分别为11、9、3、3和3。基因结构和蛋白保守基序分析表明, 谷子HAK家族具有较高的保守性, 不同Cluster的保守性依次为: Cluster III = Cluster V > Cluster II > Cluster I > Cluster IV。(2) 串联复制是SiHAKs扩增的主要原因, 15个SiHAKs位于串联重复中。(3) 171个转录因子可能结合到不同SiHAKs的启动子上, 这些转录因子包含ERF、NAC、MYB和WRKY等家族中的大量成员, 可能授予了SiHAKs对非生物胁迫多样的响应机制。(4) 基因表达聚类将SiHAKs分成3组: Group I、Group II和Group III, 多数SiHAKs在张谷和豫谷1号2个品种中的表达模式具有一致性; 不同Cluster表达水平总体表现为: Cluster III > Cluster V > Cluster II > Cluster I > Cluster IV。(5) 根系中表达水平较高的11个SiHAKs用来检测对低钾和高盐胁迫的响应。在低钾胁迫后, 8个SiHAKs的表达水平显著升高, 1个SiHAK显著降低, 2个SiHAKs变化不明显; 而高盐胁迫后, 3个SiHAKs的表达水平显著升高, 2个SiHAKs显著降低, 其余6个SiHAKs变化不明显。SiHAK15受到低钾和高盐胁迫的响应最为强烈, 其表达量分别为对照的151倍和22倍。(6) 基因表达谱的差异反映出不同Cluster间SiHAKs的功能差异。Cluster I主要在根系中表达, 可能参与谷子根系K+的吸收; Cluster II不具有组织表达特异性, 推测其参与K+的吸收、转运和生长发育等多个生物过程; Cluster III受到低钾和高盐2种胁迫的诱导, 显示出维持谷子K+/Na+平衡和抵御盐胁迫的潜在作用; Cluster IV在被检测的多个组织中几乎不表达; Cluster V不同成员对低钾和高盐胁迫的响应存在差异, 可能发生了功能分化。研究结果不仅为深入解析谷子HAK家族的功能奠定了基础, 而且为植物中钾高效利用和耐盐机制的研究提供了重要线索。
| [1] |
Zörb C, Senbayram M, Peiter E. Potassium in agriculture: status and perspectives. J Plant Physiol, 2014, 171: 656-669.
doi: 10.1016/j.jplph.2013.08.008 |
| [2] |
Lu Z, Pan Y, Hu W, Cong R, Ren T, Guo S, Lu J. The photosynthetic and structural differences between leaves and siliques of Brassica napus exposed to potassium deficiency. BMC Plant Biol, 2017, 17: 240.
doi: 10.1186/s12870-017-1201-5 |
| [3] |
Hasanuzzaman M, Bhuyan M H M B, Nahar K, Hossain M S, Mahmud J A, Hossen M S, Masud A A C, Moumita, Fujita M. Potassium: a vital regulator of plant responses and tolerance to abiotic stresses. Agronomy, 2018, 8: 31.
doi: 10.3390/agronomy8030031 |
| [4] |
Wang Y, Wu W H. Regulation of potassium transport and signaling in plants. Curr Opin Plant Biol, 2017, 39: 123-128.
doi: S1369-5266(17)30031-6 pmid: 28710919 |
| [5] |
Mostofa M G, Rahman M M, Ghosh T K, Kabir A H, Abdelrahman M, Khan M A R, Mochida K, Tran L P. Potassium in plant physiological adaptation to abiotic stresses. Plant Physiol Biochem, 2022, 186: 279-289.
doi: 10.1016/j.plaphy.2022.07.011 |
| [6] |
Johnson R, Vishwakarma K, Hossen M S, Kumar V, Shackira A M, Puthur J T, Abdi G, Sarraf M, Hasanuzzaman M. Potassium in plants: growth regulation, signaling, and environmental stress tolerance, Plant Physiol Biochem, 2022, 172: 56-69.
doi: 10.1016/j.plaphy.2022.01.001 |
| [7] |
Hussain S, Hussain S, Ali B, Ren X, Chen X, Li Q, Saqib M, Ahmad N. Recent progress in understanding salinity tolerance in plants: story of Na+/K+ balance and beyond. Plant Physiol Biochem, 2021, 160: 239-256.
doi: 10.1016/j.plaphy.2021.01.029 |
| [8] |
Gierth M, Mäser P. Potassium transporters in plants: involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett, 2007, 581: 2348-2356.
doi: 10.1016/j.febslet.2007.03.035 |
| [9] |
Li W, Xu G, Alli A, Yu L. Plant HAK/KUP/KT K+ transporters: function and regulation. Semin Cell Dev Biol, 2018, 74: 133-141.
doi: 10.1016/j.semcdb.2017.07.009 |
| [10] |
Ahn S J, Shin R, Schachtman D P. Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake. Plant Physiol, 2004, 134: 1135-1145.
doi: 10.1104/pp.103.034660 |
| [11] |
Gupta M, Qiu X, Wang L, Xie W, Zhang C, Xiong L, Lian X, Zhang Q. KT/HAK/KUP potassium transporters gene family and their whole-life cycle expression profile in rice (Oryza sativa). Mol Genet Genomics, 2008, 280: 437-452.
doi: 10.1007/s00438-008-0377-7 pmid: 18810495 |
| [12] |
Li Y, Peng L, Xie C, Shi X, Dong C, Shen Q, Xu Y. Genome-wide identification, characterization, and expression analyses of the HAK/KUP/KT potassium transporter gene family reveals their involvement in K+ deficient and abiotic stress responses in pear rootstock seedlings. Plant Growth Regul, 2018, 85: 187-198.
doi: 10.1007/s10725-018-0382-8 |
| [13] |
He C, Cui K, Duan A, Zeng Y, Zhang J. Genome-wide and molecular evolution analysis of the Poplar KT/HAK/KUP potassium transporter gene family. Ecol Evol, 2012, 2: 1996-2004.
doi: 10.1002/ece3.299 pmid: 22957200 |
| [14] |
Cheng X, Liu X, Mao W, Zhang X, Chen S, Zhan K, Bi H, Xu H. Genome-wide identification and analysis of HAK/KUP/KT potassium transporters gene family in wheat (Triticum aestivum L.). Int J Mol Sci, 2018, 19: 3969.
doi: 10.3390/ijms19123969 |
| [15] |
Zhou J, Zhou H J, Chen P, Zhang L L, Zhu J T, Li P F, Yang J, Ke Y Z, Zhou Y H, Li J N, Du H. Genome-wide survey and expression analysis of the KT/HAK/KUP family in Brassica napus and its potential roles in the response to K+ deficiency. Int J Mol Sci, 2020, 21: 9487.
doi: 10.3390/ijms21249487 |
| [16] | 许赛赛, 张博, 仲阳, 段思凡, 杨慧芹, 马玲. 马铃薯HAK/ KUP/KT基因家族鉴定与表达分析. 分子植物育种, 2021, 19: 3878-3886. |
| Xu S S, Zhang B, Zhong Y, Duan S F, Yang H Q, Ma L. Identification and expression analysis of HAK/KUP/KT gene family in potato. Mol Plant Breed, 2021, 19: 3878-3886. (in Chinese with English abstract) | |
| [17] |
Rubio F, Santa-María G E, Rodríguez-Navarro A. Cloning of Arabidopsis and barley cDNA encoding HAK potassium transporters in root and shoot cells. Physiol Plant, 2001, 109: 34-43.
doi: 10.1034/j.1399-3054.2000.100106.x |
| [18] |
Nieves-Cordones M, Ródenas R, Chavanieu A, Rivero R M, Martinez V, Gaillard I, Rubio F. Uneven HAK/KUP/KT protein diversity among angiosperms: species distribution and perspectives. Front Plant Sci, 2016, 7: 127.
doi: 10.3389/fpls.2016.00127 pmid: 26904084 |
| [19] |
Santa-María G E, Rubio F, Dubcovsky J, Rodríguez-Navarro A. The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter. Plant Cell, 1997, 9: 2281-2289.
doi: 10.1105/tpc.9.12.2281 pmid: 9437867 |
| [20] |
Gierth M, Mäser P, Schroeder J I. The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol, 2005, 137: 1105-1114.
doi: 10.1104/pp.104.057216 |
| [21] |
Bañuelos M A, Garciadeblas B, Cubero B, Rodríguez-Navarro A. Inventory and functional characterization of the HAK potassium transporters of rice. Plant Physiol, 2002, 130: 784-795.
doi: 10.1104/pp.007781 pmid: 12376644 |
| [22] |
Han M, Wu W, Wu W H, Wang Y. Potassium transporter KUP7 is involved in K+ Acquisition and translocation in Arabidopsis root under K+-limited conditions. Mol Plant, 2016, 9: 437-446.
doi: 10.1016/j.molp.2016.01.012 |
| [23] |
Okada T, Nakayama H, Shinmyo A, Yoshida K. Expression of OsHAK genes encoding potassium ion transporters in rice. Plant Biotechnol, 2008, 25: 241-245.
doi: 10.5511/plantbiotechnology.25.241 |
| [24] |
晁毛妮, 温青玉, 张志勇, 胡根海, 张金宝, 王果, 王清连. 陆地棉钾转运体基因GhHAK5的序列特征及表达分析. 作物学报, 2018, 44: 236-244.
doi: 10.3724/SP.J.1006.2018.00236 |
|
Chao M N, Wen Q Y, Zhang Z Y, Hu G H, Zhang J B, Wang G, Wang Q L. Sequence characteristics and expression analysis of potassium transporter gene GhHAK5 in upland cotton (Gossypium hirsutum L.). Acta Agron Sin, 2018, 44: 236-244. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2018.00236 |
|
| [25] |
Senn M E, Rubio F, Bañuelos M A, Rodríguez-Navarro A. Comparative functional features of plant potassium HvHAK1 and HvHAK2 transporters. J Biol Chem, 2001, 276: 44563-44569.
doi: 10.1074/jbc.M108129200 pmid: 11562376 |
| [26] |
Garciadeblas B, Benito B, Rodríguez-Navarro A.Molecular cloning and functional expression in bacteria of the potassium transporters CnHAK1 and CnHAK2of the seagrass Cymodocea nodosa. Plant Mol Biol, 2002, 50: 623-633.
pmid: 12374296 |
| [27] |
Wang X, Li J, Li F, Pan Y, Cai D, Mao D, Chen L, Luan S. Rice potassium transporter OsHAK8 mediates K+ uptake and translocation in response to low K+ stress. Front Plant Sci, 2021, 12: 730002.
doi: 10.3389/fpls.2021.730002 |
| [28] |
Osakabe Y, Arinaga N, Umezawa T, Katsura S, Nagamachi K, Tanaka H, Ohiraki H, Yamada K, Seo S U, Abo M, Yoshimura E, Shinozaki K, Yamaguchi-Shinozaki K. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell, 2013, 25: 609-624.
doi: 10.1105/tpc.112.105700 |
| [29] |
Chen G, Liu C, Gao Z, Zhang Y, Jiang H, Zhu L, Ren D, Yu L, Xu G, Qian Q. OsHAK1, a high-affinity potassium transporter, positively regulates responses to drought stress in rice. Front Plant Sci, 2017, 8: 1885.
doi: 10.3389/fpls.2017.01885 pmid: 29163608 |
| [30] |
Feng H, Tang Q, Cai J, Xu B, Xu G, Yu L. Rice OsHAK 16 functions in potassium uptake and translocation in shoot, maintaining potassium homeostasis and salt tolerance. Planta, 2019, 250: 549-561.
doi: 10.1007/s00425-019-03194-3 |
| [31] |
Zhang M, Liang X, Wang L, Cao Y, Song W, Shi J, Lai J, Jiang C. A HAK family Na+ transporter confers natural variation of salt tolerance in maize. Nat Plants, 2019, 5: 1297-1308.
doi: 10.1038/s41477-019-0565-y pmid: 31819228 |
| [32] |
Jing X, Song X, Cai S, Wang P, Lu G, Yu L, Zhang C, Wu Z. Overexpression of OsHAK5 potassium transporter enhances virus resistance in rice (Oryza sativa). Mol Plant Pathol, 2022, 23: 1107-1121.
doi: 10.1111/mpp.v23.8 |
| [33] |
刁现民, 程汝宏.十五年区试数据分析展示谷子糜子育种现状中国农业科学, 2017, 50: 4469-4474.
doi: 10.3864/j.issn.0578-1752.2017.23.001 |
| Diao X M, Cheng R H. Current breeding situation of foxtail millet and common millet in China as revealed by exploitation of 15 years regional adaptation test data. Sci Agric Sin, 2017, 50: 4469-4474. (in Chinese with English abstract) | |
| [34] | 张亚琦, 李淑文, 杜雄, 文宏达. 施钾对杂交谷子水分利用效率和产量的影响. 河北农业大学学报, 2014, 37(6): 1-6. |
| Zhang Y Q, Li S W, Du X, Wen H D. Effect of potassium fertilization on water use efficiency and yield of hybrid millet. J Hebei Agric Univ, 2014, 37(6): 1-6. (in Chinese with English abstract) | |
| [35] | 宋淑贤, 田伯红, 王建广, 赵光辉, 刘艳丽, 张立新. 不同施钾量对谷子生长及产量的影响. 辽宁农业科学, 2015, (6): 6-8. |
| Song S X, Tian B H, Wang J G, Zhao G H, Liu Y L, Zhang L X. Effect of potassium fertilization on the growth and yield of millet. Liaoning Agric Sci, 2015, (6): 6-8 (in Chinese with English abstract). | |
| [36] | 郝科星, 李娜娜, 侯富恩. 氮·磷·钾肥运筹对谷子品质与产量的影响. 安徽农业科学, 2016, 44(13): 51-55. |
| Hao K X, Li N N, Hou H E. Effect of N, P, K Fertilizer management on the quality and yield of millet. J Anhui Agric Sci, 2016, 44(13): 51-55. (in Chinese with English abstract) | |
| [37] |
Nadeem F, Ahmad Z, Hassan M U, Wang R, Diao X, Li X. Adaptation of foxtail millet (Setaria italica L.) to abiotic stresses: a special perspective of responses to nitrogen and phosphate limitations. Front Plant Sci, 2020, 11: 187.
doi: 10.3389/fpls.2020.00187 |
| [38] |
陈二影, 王润丰, 秦岭, 杨延兵, 黎飞飞, 张华文, 王海莲, 刘宾, 孔清华, 管延安. 谷子芽期耐盐碱综合鉴定及评价. 作物学报, 2020, 46: 1591-1604.
doi: 10.3724/SP.J.1006.2020.04064 |
| Chen E Y, Wang R F, Qin L, Yang Y B, Li F F, Zhang H W, Wang H L, Liu B, Kong Q H, Guan Y A. Comprehensive identification and evaluation of foxtail millet for saline-alkaline tolerance during germination. Acta Agron Sin, 2020, 46: 1591-1604. (in Chinese with English abstract) | |
| [39] |
Sato Y, Nanatani K, Hamamoto S, Shimizu M, Takahashi M, Tabuchi-Kobayashi M, Mizutani A, Schroeder J I, Souma S, Uozumi N. Defining membrane spanning domains and crucial membrane-localized acidic amino acid residues for K+ transport of a Kup/HAK/KT-type Escherichia coli potassium transporter. J Biochem, 2014, 155: 315-323.
doi: 10.1093/jb/mvu007 |
| [40] |
Cannon S B, Mitra A, Baumgarten A, Young N D, May G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol, 2004, 4: 10.
pmid: 15171794 |
| [41] | 宋毓峰, 张良, 董连红, 靳义荣, 史素娟, 白岩, 刘朝科, 冯广林, 冯祥国, 王倩, 刘好宝. 植物KUP/HAK/KT家族钾转运体研究进展. 中国农业科技导报, 2013, 15(6): 92-98. |
| Song Y F, Zhang L, Dong L H, Jin Y R, Shi S J, Bai Y, Liu C K, Feng G L, Feng X G, Wang Q, Liu H B. Research progress on KUP/HAK/KT potassium transporter family in plant. J Agric Sci Technol, 2013, 15(6): 92-98. (in Chinese with English abstract) | |
| [42] | 柴薇薇, 王文颖, 崔彦农, 王锁民. 植物钾转运蛋白KUP/HAK/KT家族研究进展. 植物生理学报, 2019, 55: 1747-1761. |
| Chai W W, Wang W Y, Cui Y N, Wang S M. Research progress of function on KUP/HAK/KT family in plants. Plant Physiol J, 2019, 55: 1747-1761. (in Chinese with English abstract) | |
| [43] |
Feng X, Liu W, Qiu C W, Zeng F, Wang Y, Zhang G, Chen Z H, Wu F. HvAKT2 and HvHAK1 confer drought tolerance in barley through enhanced leaf mesophyll H+ homoeostasis. Plant Biotechnol J, 2020, 18: 1683-1696.
doi: 10.1111/pbi.13332 pmid: 31917885 |
| [44] |
Kim E J, Kwak J M, Schroeder U J I. AtKUP1: an Arabidopsis gene encoding high-affinity potassium transport activity. Plant Cell, 1998, 10: 51-62.
pmid: 9477571 |
| [45] |
Uchiyama M, Fudaki R, Kobayashi T, Adachi Y, Ukai Y, Yoshihara T, Shimada H. Rice OsHAK5 is a major potassium transporter that functions in potassium uptake with high specificity but contributes less to cesium uptake. Biosci Biotechnol Biochem, 2022, 86: 1599-1604.
doi: 10.1093/bbb/zbac152 |
| [46] |
Yang T, Zhang S, Hu Y, Wu F, Hu Q, Chen G, Cai J, Wu T, Moran N, Yu L, Xu G. The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol, 2014, 166: 945-959.
doi: 10.1104/pp.114.246520 pmid: 25157029 |
| [47] |
Maathuis F. The role of monovalent cation transporters in plant responses to salinity. J Exp Bot, 2006, 57: 1137-1147.
doi: 10.1093/jxb/erj001 pmid: 16263900 |
| [48] |
Zhang L, Sun X, Li Y, Luo X, Song S, Chen Y, Wang X, Mao D, Chen L, Luan S. Rice Na+-permeable transporter OsHAK12 mediates shoots Na+ exclusion in response to salt stress. Front Plant Sci, 2021, 12: 771746.
doi: 10.3389/fpls.2021.771746 |
| [1] | 徐扬, 张岱, 康涛, 温赛群, 张冠初, 丁红, 郭庆, 秦斐斐, 戴良香, 张智猛. 盐胁迫对花生幼苗离子动态及耐盐基因表达的影响[J]. 作物学报, 2023, 49(9): 2373-2384. |
| [2] | 文利超, 熊涛, 邓智超, 刘涛, 郭存, 李伟, 郭永峰. 烟草转录因子NtNAC080在非生物胁迫下的表达分析及功能鉴定[J]. 作物学报, 2023, 49(8): 2171-2182. |
| [3] | 万夷曼, 肖圣慧, 白依超, 范佳音, 王琰, 吴长艾. 谷子毛状根诱导方法的建立与优化[J]. 作物学报, 2023, 49(7): 1758-1768. |
| [4] | 丁洪艳, 冯晓溪, 汪柏宇, 张积森. 甘蔗割手密种LRRII-RLK基因家族演化和表达分析[J]. 作物学报, 2023, 49(7): 1769-1784. |
| [5] | 梅玉琴, 刘意, 王崇, 雷剑, 朱国鹏, 杨新笋. 甘薯PHB基因家族的全基因组鉴定和表达分析[J]. 作物学报, 2023, 49(6): 1715-1725. |
| [6] | 刘佳, 邹晓悦, 马继芳, 王永芳, 董志平, 李志勇, 白辉. 谷子MAPK家族成员的鉴定及其对生物胁迫的响应分析[J]. 作物学报, 2023, 49(6): 1480-1495. |
| [7] | 贾玉库, 高宏欢, 冯健超, 郝紫瑞, 王晨阳, 谢迎新, 郭天财, 马冬云. 小麦G2-like转录因子家族基因鉴定与表达模式分析[J]. 作物学报, 2023, 49(5): 1410-1425. |
| [8] | 张小红, 彭琼, 鄢铮. 盐胁迫下不同甘薯品种的转录组测序分析[J]. 作物学报, 2023, 49(5): 1432-1444. |
| [9] | 孙全喜, 苑翠玲, 牟艺菲, 闫彩霞, 赵小波, 王娟, 王奇, 孙慧, 李春娟, 单世华. 花生SWEET基因全基因组鉴定及表达分析[J]. 作物学报, 2023, 49(4): 938-954. |
| [10] | 齐燕妮, 李闻娟, 赵丽蓉, 李雯, 王利民, 谢亚萍, 赵玮, 党照, 张建平. 亚麻生氰糖苷合成关键酶CYP79基因家族的鉴定及表达分析[J]. 作物学报, 2023, 49(3): 687-702. |
| [11] | 张文宣, 梁晓梅, 戴成, 文静, 易斌, 涂金星, 沈金雄, 傅廷栋, 马朝芝. 利用CRISPR/Cas9技术突变BnaMPK6基因降低甘蓝型油菜的耐盐性[J]. 作物学报, 2023, 49(2): 321-331. |
| [12] | 王蓉, 陈小红, 王倩, 刘少雄, 陆平, 刁现民, 刘敏轩, 王瑞云. 中国谷子名米品种遗传多样性与亲缘关系研究[J]. 作物学报, 2022, 48(8): 1914-1925. |
| [13] | 王沙沙, 黄超, 汪庆昌, 晁岳恩, 陈锋, 孙建国, 宋晓. 小麦籽粒大小相关基因TaGS2克隆及功能分析[J]. 作物学报, 2022, 48(8): 1926-1937. |
| [14] | 郭家鑫, 鲁晓宇, 陶一凡, 郭慧娟, 闵伟. 棉花在盐碱胁迫下代谢产物及通路的分析[J]. 作物学报, 2022, 48(8): 2100-2114. |
| [15] | 韩尚玲, 霍轶琼, 李辉, 韩华蕊, 侯思宇, 孙朝霞, 韩渊怀, 李红英. 基于WGCNA发掘谷子穗部类黄酮合成途径调控关键基因[J]. 作物学报, 2022, 48(7): 1645-1657. |
|