欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (4): 944-956.doi: 10.3724/SP.J.1006.2024.34141
张慧( ), 张欣雨, 袁旭, 陈伟达, 杨婷()
), 张欣雨, 袁旭, 陈伟达, 杨婷()
ZHANG Hui(), ZHANG Xin-Yu, YUAN Xu, CHEN Wei-Da, YANG Ting()
摘要:
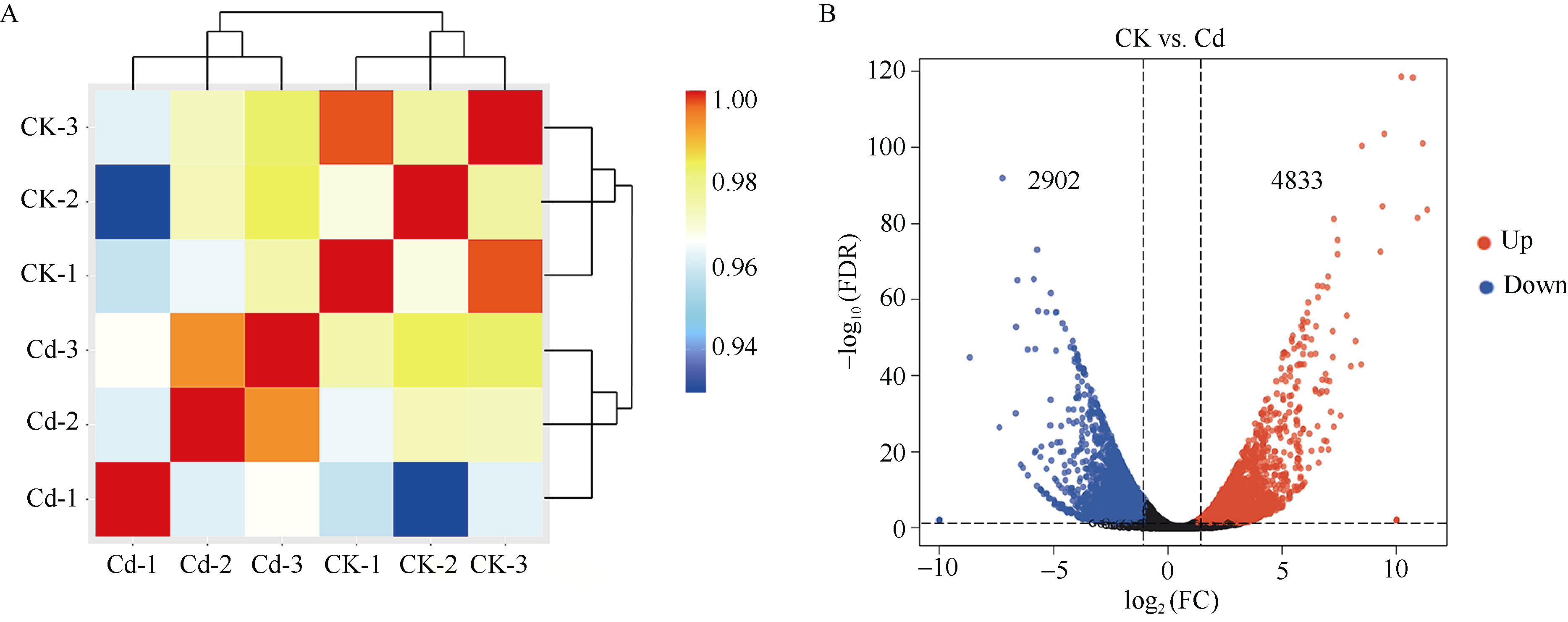
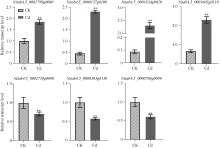
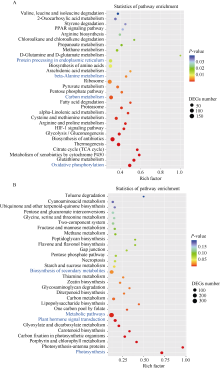
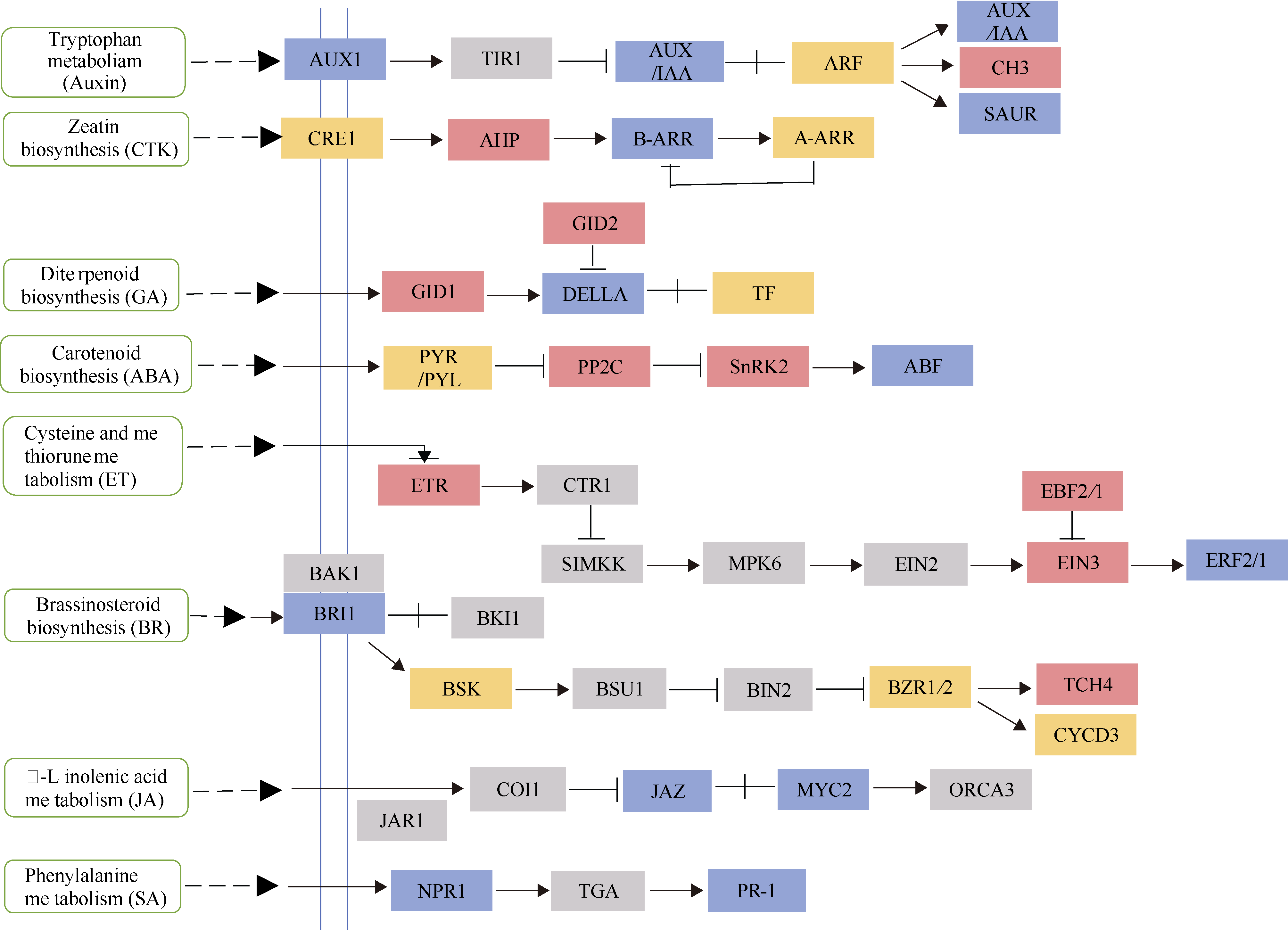
烟草具有超富集镉的能力, 严重降低烟叶品质, 影响其经济价值。为了阐释烟草响应镉胁迫的分子机制, 本研究采集了镉浓度为0和500 μmol L-1培养条件下的烟草叶片进行转录组测序。共获得76.94 Gb有效数据(Clean data), Q30 碱基百分比均达到95.43%以上; 在镉胁迫的烟草叶片中, 共筛选出7735个差异表达基因, 其中4833个基因表达上调, 2902个基因表达下调, 并通过qRT-PCR分析验证了转录组数据的可靠性。对差异转录本进行GO和KEGG富集分析, GO注释表明差异基因涉及代谢过程、应激反应、细胞结构体、催化活性和转录调节活性等; KEGG富集分析表明上调差异基因主要富集在氨基酸的生物合成、碳代谢、氧化磷酸化和柠檬酸循环等通路, 下调差异基因则主要富集在光合作用、次生代谢产物的生物合成、代谢途径和植物激素信号转导途径。进一步分析植物激素信号转导通路发现, 共有8条植物激素途径以不同的表达方式参与烟草对镉胁迫的响应。激素喷施烟草的实验结果表明, 叶片通过调控赤霉素、油菜素内酯和茉莉酸途径以应对镉胁迫; 拟南芥激素信号缺失突变体验证实验表明赤霉素、油菜素内酯、茉莉酸和乙烯途径均响应镉胁迫。综上所述, 本文以转录组分析探究了烟草叶片响应镉胁迫的调控网络, 以期为提高作物抗逆性的遗传改良提供理论依据。
| [1] | 刘娟, 张乃明, 于泓, 张靖宇, 李芳艳, 于畅, 杜红蝶. 重金属污染对水稻土微生物及酶活性影响研究进展. 土壤, 2021, 53: 1152-1159. |
| Liu J, Zhang L M, Yu H, Zhang J Y, Li F Y, Yu C, Du H D. Effects of heavy metal pollution on microorganism and enzyme activity in paddy soil: a review. Soils, 2021, 53: 1152-1159. (in Chinese with English abstract) | |
| [2] | 苏芸芸. 乙酰胆碱调节烟草Cd胁迫响应的生理机制. 西北农林科技大学博士学位论文, 陕西杨凌, 2021. |
| Su Y Y. The Physiological Response Mechanism of Acetylcholine Regulating Cadmium Stress in Tobacco. PhD Dissertation of Northwest A&F University, Yangling, Shaanxi, China, 2021. (in Chinese with English abstract) | |
| [3] |
Järup L, Akesson A. Current status of cadmium as an environmental health problem. Toxicol Appl Pharm, 2009, 238: 201-208.
doi: 10.1016/j.taap.2009.04.020 |
| [4] |
Shifaw E. Review of heavy metals pollution in China in agricultural and urban soils. J Health Pollut, 2018, 8: 180607.
doi: 10.5696/2156-9614-8.18.180607 |
| [5] |
Li D, He T, Saleem M, He G. Metalloprotein-specific or critical amino acid residues: perspectives on plant-precise detoxification and recognition mechanisms under cadmium stress. Int J Mol Sci, 2022, 23: 1734.
doi: 10.3390/ijms23031734 |
| [6] |
Yang Z, Yang F, Liu J L, Wu H T, Yang H, Shi Y, Jie L, Zhang Y F, Luo Y R, Chen K M. Heavy metal transporters: functional mechanisms regulation and application in phytoremediation. Sci Total Environ, 2022, 809: 151099.
doi: 10.1016/j.scitotenv.2021.151099 |
| [7] |
Haider F U, Cai L Q, Coulter J A, Cheema S A, Wu J, Zhang R Z, Ma W J, Farooq M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotox Environ Safe, 2021, 211: 111887.
doi: 10.1016/j.ecoenv.2020.111887 |
| [8] |
de Araújo R P, de Almeida A A F, Pereira L S, Mangabeira P A O, Souza J O, Pirovani C P, Ahnert D, Baligar V C. Photosynthetic, antioxidative, molecular and ultrastructural responses of young cacao plants to Cd toxicity in the soil. Ecotox Environ Safe, 2017, 144: 148-157.
doi: S0147-6513(17)30331-7 pmid: 28614756 |
| [9] |
Dong X X, Yang F, Yang S P, Yan C Z. Subcellular distribution and tolerance of cadmium in Canna indica L. Ecotox Environ Safe, 2019, 185: 109692.
doi: 10.1016/j.ecoenv.2019.109692 |
| [10] |
Guo L, Chen A, He N, Yang D, Liu M D. Exogenous silicon alleviates cadmium toxicity in rice seedlings in relation to Cd distribution and ultrastructure changes. J Soil Sediment, 2018, 18: 1691-1700.
doi: 10.1007/s11368-017-1902-2 |
| [11] |
Raza A, Habib M, Kakavand S N, Zahid Z, Zahra N, Sharif R. Hasanuzzaman M. Phytoremediation of cadmium: physiological, biochemical, and molecular mechanisms. Biology, 2020, 9: 177.
doi: 10.3390/biology9070177 |
| [12] | Wang B, Wei H, Xue Z, Zhang W H. Gibberellins regulate iron deficiency response by influencing iron transport and translocation in rice seedlings (Oryza sativa). Ann Bot, 2017, 119: 945-956. |
| [13] |
Kumari A, Das P, Parida A K, Agarwal P K. Proteomics, metabolomics, and ionomics perspectives of salinity tolerance in halophytes. Front Plant Sci, 2015, 6: 537.
doi: 10.3389/fpls.2015.00537 pmid: 26284080 |
| [14] |
Kim J M, To T K, Matsui A, Tanoi K, Kobayashi N I, Matsuda F, Habu Y, Ogawa D, Sakamoto T, Matsunaga S, Bashir1 K, Rasheed S, Ando M, Takeda H, Kawaura K, Kusano M, Fukushima A, Endo T A, Kuromori T, Ishida J, Morosawa T, Tanaka M, Torii C, Takebayashi Y, Sakakibara H, Ogihara Y, Saito K, Shinozaki K, Devoto A, Seki M. Acetate-mediated novel survival strategy against drought in plants. Nat Plants, 2017, 3: 17097.
doi: 10.1038/nplants.2017.97 |
| [15] |
Abozeid A, Ying Z, Lin Y, Liu J, Zhang Z, Tang Z. Ethylene improves root system development under cadmium stress by modulating superoxide anion concentration in Arabidopsis thaliana. Front Plant Sci, 2017, 8: 253.
doi: 10.3389/fpls.2017.00253 pmid: 28286514 |
| [16] |
Borgo L, Rabêlo F H S, Budzinski I G F, Cataldi T R, Ramires T G, Schaker P D C, Ribas A F, Labate C A, Lavres J, Cuypers A, Azevedo R A. Proline exogenously supplied or endogenously overproduced induces different nutritional, metabolic, and antioxidative responses in transgenic tobacco exposed to cadmium. J Plant Growth Regul, 2022, 41: 2846-2868.
doi: 10.1007/s00344-021-10480-6 |
| [17] |
Xu J, Wang W Y, Sun J H, Zhang Y, Ge Q, Du L G, Yin H X, Liu X J. Involvement of auxin and nitric oxide in plant Cd-stress responses. Plant Soil, 2011, 346: 107-119.
doi: 10.1007/s11104-011-0800-4 |
| [18] |
Rosén K, Eriksson J, Vinichuk M. Uptake and translocation of 109Cd and stable Cd within tobacco plants (Nicotiana sylvestris). J Environ Radioac, 2012, 113: 16-20.
doi: 10.1016/j.jenvrad.2012.04.008 |
| [19] |
Liu H W, Wang H Y, Ma Y B, Wang H H, Shi Y. Role of transpiration and metabolism in translocation and accumulation of cadmium in tobacco plants (Nicotiana tabacum L.). Chemosphere, 2016, 144: 1960-1965.
doi: 10.1016/j.chemosphere.2015.10.093 |
| [20] |
Bush P G, Mayhew T M, Abramovich D R, Aggett P J, Burke M D, Page K R. A quantitative study on the effects of maternal smoking on placental morphology and cadmium concentration. Placenta, 2000, 21: 247-256.
pmid: 10736249 |
| [21] |
Li H Q, Wallin M, Barregard L, Sallsten G, Lundh T, Ohlsson C, Mellström D, Andersson E M. Smoking-induced risk of osteoporosis is partly mediated by cadmium from tobacco smoke: the MrOS Sweden Study. J Bone Miner Res, 2020, 35: 1424-1429.
doi: 10.1002/jbmr.4014 pmid: 32191351 |
| [22] |
Regassa G, Chandravanshi B S. Levels of heavy metals in the raw and processed Ethiopian tobacco leaves. SpringerPlus, 2016, 5: 232.
doi: 10.1186/s40064-016-1770-z pmid: 27026926 |
| [23] |
Wang X K, Shi M, Hao P F, Zheng W T, Cao F B. Alleviation of cadmium toxicity by potassium supplementation involves various physiological and biochemical features in Nicotiana tabacum L. Acta Physiol Plant, 2017, 39: 132.
doi: 10.1007/s11738-017-2424-7 |
| [24] |
Edwards K D, Fernandez-Pozo N, Drake-Stowe K, Humphry M, Evans A D, Bombarely A, Allen F, Hurst R, White B, Kernodle S P, Bromley J R, Sanchez-Tamburrino J P, Lewis R S, Mueller L A. A reference genome for Nicotiana tabacum enables map-based cloning of homeologous loci implicated in nitrogen utilization efficiency. BMC Genomics, 2017, 18: 448.
doi: 10.1186/s12864-017-3791-6 pmid: 28625162 |
| [25] |
Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25: 402-408.
doi: 10.1006/meth.2001.1262 |
| [26] |
Eissa M A. Phytoextraction mechanism of Cd by Atriplex lentiform is using some mobilizing agents. Ecol Eng, 2017, 108: 220-226.
doi: 10.1016/j.ecoleng.2017.08.025 |
| [27] | Hasan M K, Ahammed G J, Yin L L, Shi K, Xia X J, Zhou Y H, Zhou J. Melatonin mitigates cadmium phytotoxicity through modulation of phytochelatins biosynthesis, vacuolar sequestration, and antioxidant potential in Solanum lycopersicum L. Front Plant Sci, 2015, 6: 601. |
| [28] | Cao Z W, Fang Y L, Lu Y H, Tan D X, Du C H, Li Y M, Ma Q L, Yu J M, Chen M Y, Zhou C, Pei L P, Zhang L, Ran H Y, He M D, Yu Z P, Zhou Z. Melatonin alleviates cadmium induced liver injury by inhibiting the TXNIP-NLRP3 inflammasome. J Pineal Res, 2017, 62: 12389. |
| [29] |
Khanna K, Kohli S K, Ohri P, Bhardwaj R, Ahmad P. Agroecotoxicological aspect of Cd in soil-plant system: uptake, translocation and amelioration strategies. Environ Sci Pollut Res, 2022, 29: 30908-30934.
doi: 10.1007/s11356-021-18232-5 |
| [30] | Soniya E V, Srinivasan A, Menon A, Kattupalli D. Transcriptomics in Response of Biotic Stress in Plants. Academic Press, San Diego, CA, USA, 2023. pp 285-303. |
| [31] |
Ghorbel M, Brini F, Sharma A, Landi M. Role of jasmonic acid in plants: the molecular point of view. Plant Cell Rep, 2021, 40: 1471-1494.
doi: 10.1007/s00299-021-02687-4 pmid: 33821356 |
| [32] | Abozeid A, Ying Z J, Lin Y C, Liu J, Zhang Z H, Tang Z H. Ethylene improves root system development under cadmium stress by modulating superoxide anion concentration in Arabidopsis thaliana. Plant Sci, 2017, 8: 253. |
| [33] |
Gallego S M, Pena L B, Barcia R A, Azpilicueta C E, Iannone M F, Rosales E P, Zawoznik M S, Groppa M D, Benavides M P. Unravelling cadmium toxicity and tolerance in plants: insight into regulatory mechanisms. Environ Exp Bot, 2012, 83: 33-46.
doi: 10.1016/j.envexpbot.2012.04.006 |
| [34] |
De Carvalho C C C R, Caramujo M J. The various roles of fatty acids. Molecules, 2018, 23: 2583.
doi: 10.3390/molecules23102583 |
| [35] |
Li C Y, Hong Y, Sun J H, Wang G P, Zhou H N, Xu L T, Wang L, Xu G Y. Temporal transcriptome analysis reveals several key pathways involve in cadmium stress response in Nicotiana tabacum L. Front Plant Sci, 2023, 14: 1143349.
doi: 10.3389/fpls.2023.1143349 |
| [36] | 王俊宇, 王晓东, 马元丹, 付璐成, 周欢欢, 王斌, 张汝敏, 高燕. ‛波叶金桂’对干旱和高温胁迫的生理生态响应. 植物生态学报, 2018, 42: 681-691. |
| Wang J Y, Wang X D, Ma Y D, Fu L C, Zhou H H, Wang P, Zhang R M, Gao Y. Physiological and ecological responses to drought and heat stresses in Osmanthus fragrans ‘Boyejingui’. J Plant Ecol, 2018, 42: 681-691. (in Chinese with English abstract) | |
| [37] |
Waadt R, Seller C A, Hsu P K, Takahashi Y, Munemasa S, Schroede J. Plant hormone regulation of abiotic stress responses. Nat Rev Mol Cell Biol, 2022, 23: 680-694.
doi: 10.1038/s41580-022-00479-6 |
| [38] |
Yosefi A, Mozafari A, Javadi T. Jasmonic acid improved in vitro strawberry (Fragaria × ananassa Duch.) resistance to PEG- induced water stress. Plant Cell Tissue Organ Cult, 2020, 142: 549-558.
doi: 10.1007/s11240-020-01880-9 |
| [39] |
Liu J, Shu D F, Tan Z L, Ma M, Gou N, Gao S, Duan G Y, Kuai B K, Hu Y X, Li S P, Cui D Y. The Arabidopsis IDD14 transcription factor interacts with bZIP-type ABFs/AREBs and cooperatively regulates ABA-mediated drought tolerance. New Phytol, 2022, 236: 929-942.
doi: 10.1111/nph.v236.3 |
| [40] | Kaya C, Tuna A L, Yokaş I, Ashraf M, Ozturk M, Athar H R. The Role of Plant Hormones in Plants under Salinity Stress. In: Salinity and Water Stress. Dordrecht: Springer Netherlands, 2009. pp 45-50. |
| [41] |
Srivastava S, Srivastava A K, Suprasanna P, D’Souza S F. Identification and profiling of arsenic stress-induced miRNAs in Brassica juncea. J Exp Bot, 2013, 64: 303-315.
doi: 10.1093/jxb/ers333 pmid: 23162117 |
| [42] |
Hac-Wydro K, Sroka A, Jablonaka K. The impact of auxins used in assisted phytoextraction of metals from the contaminated environment on thea alterations caused by lead (II) ions in the organization of model lipid membranes. Colloid Surface B, 2016, 143: 124-130.
doi: 10.1016/j.colsurfb.2016.03.018 |
| [43] |
Chen H F, Zhang Q, Lv W, Yu X Y, Zhang Z H. Ethylene positively regulates Cd tolerance via reactive oxygen species scavenging and apoplastic transport barrier formation in rice. Environ Pollut, 2022, 302: 119063.
doi: 10.1016/j.envpol.2022.119063 |
| [44] |
Hayat S, Ali B, Hasan S A, Ahmad A. Brassinosteroid enhanced the level of antioxidants under cadmium stress in Brassica juncea. Environ Exp Bot, 2007, 60: 33-41.
doi: 10.1016/j.envexpbot.2006.06.002 |
| [45] |
Chen H, Yang R X, Zhang X, Chen Y H, Xia Y, Xu X M. Foliar application of gibberellin inhibits the cadmium uptake and xylem transport in lettuce (Lactuca sativa L.). Sci Hortic, 2021, 288: 110410.
doi: 10.1016/j.scienta.2021.110410 |
| [46] |
Lu Q Y, Chen S M, Li Y Y, Zheng F H, He B, Gu M H. Exogenous abscisic acid (ABA) promotes cadmium (Cd) accumulation in Sedum alfredii Hance by regulating the expression of Cd stress response genes. Environ Sci Pollut Res, 2020, 27: 8719-8731.
doi: 10.1007/s11356-019-07512-w |
| [1] | 曹松, 姚敏, 任睿, 贾元, 向星汝, 李文, 何昕, 刘忠松, 官春云, 钱论文, 熊兴华. 转录组结合区域关联分析挖掘油菜含油量积累的候选基因[J]. 作物学报, 2024, 50(5): 1136-1146. |
| [2] | 宋松泉, 唐翠芳, 雷华平, 姜孝成, 王伟青, 程红焱. 种子休眠与萌发调控的研究进展[J]. 作物学报, 2024, 50(1): 1-15. |
| [3] | 李明月, 张文婷, 李阳, 张保龙, 杨立明, 王金彦. 小肽Ospep5对水稻耐镉性的影响[J]. 作物学报, 2024, 50(1): 67-75. |
| [4] | 王菲菲, 张胜忠, 胡晓辉, 崔凤高, 钟文, 赵立波, 张天雨, 郭进涛, 于豪谅, 苗华荣, 陈静. 比较转录组分析花生种子休眠调控网络[J]. 作物学报, 2023, 49(9): 2446-2461. |
| [5] | 李凌雨, 周琦锐, 李洋, 张安民, 王贝贝, 马尚宇, 樊永惠, 黄正来, 张文静. 外源6-BA调控孕穗期低温后小麦幼穗发育的转录组分析[J]. 作物学报, 2023, 49(7): 1808-1817. |
| [6] | 赵晓鑫, 黄烁淇, 谭文勃, 兴旺, 刘大丽. 甜菜HIPPs基因家族鉴定与镉胁迫下的表达分析[J]. 作物学报, 2023, 49(12): 3302-3314. |
| [7] | 丁孟丽, 王茹茵, 施栋晟, 李莹博, 雷洁, 陈洪宇, 申清文, 王桂凤. 玉米小籽粒突变体mn-Mu的基因克隆与转录组分析[J]. 作物学报, 2023, 49(11): 3122-3130. |
| [8] | 朱继杰, 王士杰, 赵红霞, 贾晓昀, 李妙, 王国印. 田间条件下不同棉花品种叶片响应化学脱叶剂噻苯隆的转录组分析[J]. 作物学报, 2023, 49(10): 2705-2716. |
| [9] | 周静远, 孔祥强, 张艳军, 李雪源, 张冬梅, 董合忠. 基于种子萌发出苗过程中弯钩建成和下胚轴生长的棉花出苗壮苗机制与技术[J]. 作物学报, 2022, 48(5): 1051-1058. |
| [10] | 曲梦雪, 宋杰, 孙菁, 胡旦旦, 王洪章, 任昊, 赵斌, 张吉旺, 任佰朝, 刘鹏. 镉胁迫对不同耐镉型玉米品种苗期根系生长的影响[J]. 作物学报, 2022, 48(11): 2945-2952. |
| [11] | 李玲红, 张哲, 陈永明, 尤明山, 倪中福, 邢界文. 普通小麦颖壳蜡质缺失突变体glossy1的转录组分析[J]. 作物学报, 2022, 48(1): 48-62. |
| [12] | 卢海, 李增强, 唐美琼, 罗登杰, 曹珊, 岳娇, 胡亚丽, 黄震, 陈涛, 陈鹏. 红麻DNA甲基化响应镉胁迫及甲基化差异基因的表达分析[J]. 作物学报, 2021, 47(12): 2324-2334. |
| [13] | 张云, 王丹媚, 王孝源, 任晴雯, 唐可, 张丽宇, 吴玉环, 刘鹏. 外源茉莉酸对菊芋镉胁迫下光合特性及镉积累的影响[J]. 作物学报, 2021, 47(12): 2490-2500. |
| [14] | 潘丽娟,陈娜,陈明娜,王通,王冕,陈静,杨珍,万勇善,禹山林,迟晓元,刘风珍. 花生AhPEPC1基因抑制表达的转基因后代转录组分析[J]. 作物学报, 2019, 45(7): 993-1001. |
| [15] | 彭振,何守朴,龚文芳,潘兆娥,贾银华,卢艳丽,杜雄明. 陆地棉幼苗NaCl胁迫下转录因子表达的转录组学分析[J]. 作物学报, 2017, 43(03): 354-370. |
|