欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (12): 2008-2016.doi: 10.3724/SP.J.1006.2020.03022
• 研究简报 • 上一篇
杨梦婷1,2( ), 张春2(), 王作平2, 邹华文1,*(), 吴忠义2,*()
), 张春2(), 王作平2, 邹华文1,*(), 吴忠义2,*()
Meng-Ting YANG1,2(), Chun ZHANG2(), Zuo-Ping WANG2, Hua-Wen ZOU1,*(), Zhong-Yi WU2,*()
摘要:
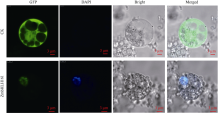
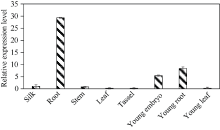
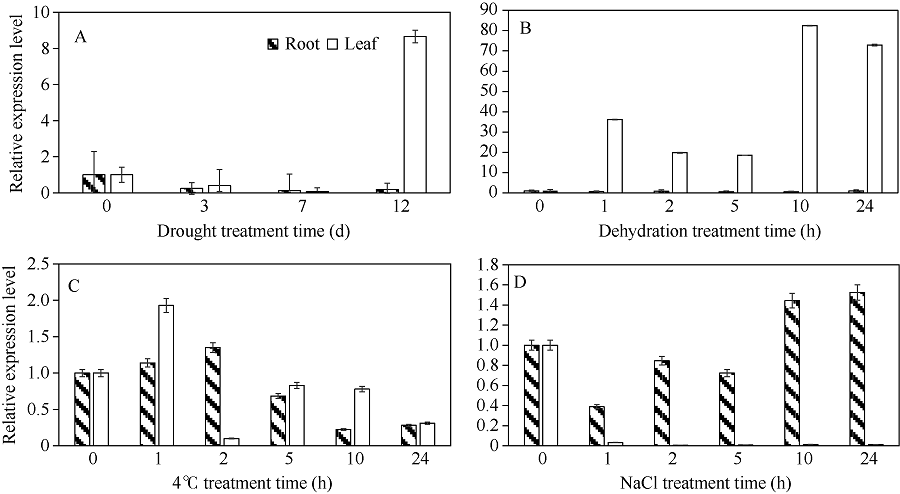
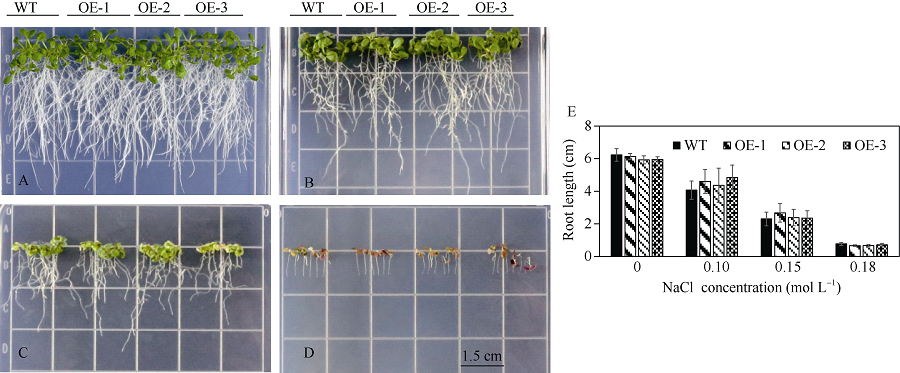
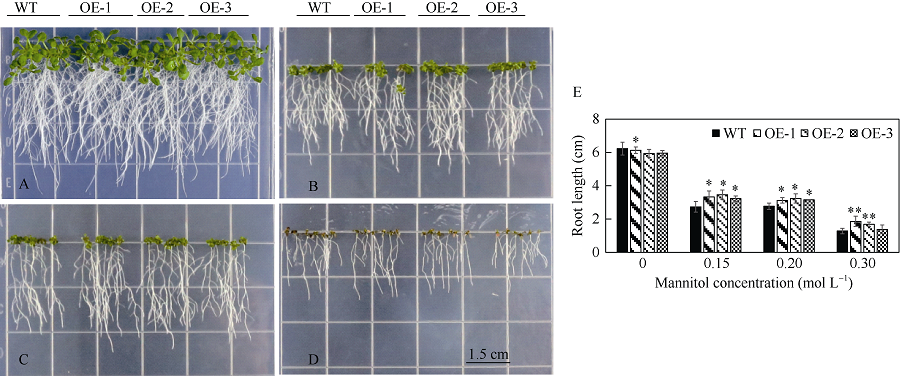
bHLH转录因子是植物第二大转录因子家族, 在调节植物生长发育、信号转导和逆境胁迫响应等方面发挥着重要的作用。为了研究玉米bHLH家族基因在逆境胁迫响应中的功能, 本研究从玉米根组织中克隆了ZmbHLH161 (AC: NC_AQK75074)基因。生物信息学分析表明: 该基因包含3个外显子, cDNA全长1460 bp, 编码序列全长1059 bp, 编码352个氨基酸; 在玉米基因组中以单拷贝形式存在, 功能未知; ZmbHLH161蛋白分子量为37.1 kD, 理论等电点为6.10, 具有bHLH转录因子家族特有的保守结构域, 但不具跨膜结构, 无信号肽, 为亲水性蛋白, 蛋白二级结构无规则卷曲所占比例最大, 为42.05%。玉米原生质体瞬时表达试验表明, ZmbHLH161定位在细胞核内。实时荧光定量PCR (qPCR)分析表明, 正常生长条件下, ZmbHLH161主要在根系和幼胚中表达; 在脱水和干旱处理下, ZmbHLH161在玉米苗期叶片中上调表达。转基因异源表达ZmbHLH161拟南芥株系经不同浓度NaCl处理后, 其根长与野生型差异不显著, 而不同浓度甘露醇处理后其根长优于野生型。由此推测ZmbHLH161基因可能参与玉米对渗透胁迫应答。
| [1] | 刘天金, 王玉玺, 宁明宇, 靖飞, 董晓霞, 王志敏, 刘春青 . 我国玉米种业转型升级的路径与策略探讨. 中国种业, 2018, ( 2):1-7. |
| Liu T J, Wang Y X, Ning M Y, Jing F, Dong X J, Wang Z M, Liu C Q . Discussion on the path and strategy of transformation and upgrading of China’s maize seed industry. China Seed Ind, 2018, ( 2):1-7 (in Chinese). | |
| [2] | 余爱丽, 赵晋锋, 王高鸿, 杜艳伟, 李颜方, 张正 . 玉米ZmSAMS1基因在盐、干旱等逆境胁迫下的表达分析. 玉米科学, 2016,24(3):31-35. |
| Yu A L, Zhao J F, Wang G H, Du Y W, Li Y F, Zhang Z . Expression analysis of ZmSAMS1 gene under salt, drought and other stress. J Maize Sci, 2016,24(3):31-35 (in Chinese with English abstract). | |
| [3] | 张艳馥, 沙伟 . 转录因子概述. 生物学教学, 2009, 34, ( 10):7-8. |
| Zhang Y F, Sha W . Overview of transcription factors. Biol Teach, 2009, 34, ( 10):7-8 (in Chinese). | |
| [4] |
Murre C, McCaw P S, Baltimore D . A new DNA binding and dimerizing motif in immunoglobulin enhancer binding, daugtherless, MyoD, and MYC proteins. Cell, 1989,56:777-783.
doi: 10.1016/0092-8674(89)90682-x pmid: 2493990 |
| [5] |
Nuno P, Liam D . Origin and diversification of basic- helix-loop-helix proteins in plants. Mol Biol Evol, 2010,27:862-874.
doi: 10.1093/molbev/msp288 pmid: 19942615 |
| [6] |
Atchley W R, Fitch W M . A natural classification of the basic helix-loop-helix class of transcription factors. Proc Natl Acad Sci USA, 1997,94:5172-5176.
doi: 10.1073/pnas.94.10.5172 pmid: 9144210 |
| [7] | 罗赛男, 杨国顺, 石雪晖, 卢向阳, 徐萍 . 转录因子在植物抗逆性上的应用研究. 湖南农业大学学报(自然科学版), 2005,31:219-223. |
| Luo S N, Yang G S, Shi X H, Lu X Y, Xu P . On the application of transcription factors to plant stress resistance. J Hunan Agric Univ (Nat Sci Edn), 2005,31:219-223 (in Chinese with English abstract). | |
| [8] |
Anna-Marie S, Sandra K, Ulrike S, Unte P H, Koen D, Heinz S . The Arabidopsis ABORTED MICROSPORES(AMS) gene encodes a MYC class transcription factor. Plant J, 2003,33:413-423.
doi: 10.1046/j.1365-313x.2003.01644.x pmid: 12535353 |
| [9] |
Dany H, Hidenori S . Antagonistic actions of HLH/bHLH proteins are involved in grain length and weight in rice. PLoS One, 2012,7:e31325.
doi: 10.1371/journal.pone.0031325 pmid: 22363621 |
| [10] |
Ikeda M, Mitsuda N, Ohme-Takagi M . ATBS1 INTERACTING FACTORs negatively regulate Arabidopsis cell elongation in the triantagonistic bHLH system. Plant Signal Behav, 2013,8:e23448.
doi: 10.4161/psb.23448 pmid: 23333962 |
| [11] |
Endo T, Fujii H, Sugiyama A, Nakano M, Nakajima N, Ikoma Y, Omura M . Overexpression of a citrus basic helix-loop-helix transcription factor ( CubHLH1), which is homologous to Arabidopsis activation-tagged bri1 suppressor 1 interacting factor genes, modulates carotenoid metabolism in transgenic tomato. Plant Sci, 2016,243:35-48.
doi: 10.1016/j.plantsci.2015.11.005 pmid: 26795149 |
| [12] |
Danielle M F, Jennifer N, Takamichi M, Julin N M, José A, Joseph R E, Masaki F, Joanne C . Three redundant brassinosteroid early response genes encode putative bHLH transcription factors required for normal growth. Genetics, 2002,162:1445-1456.
pmid: 12454087 |
| [13] |
Ni M, Tepperman J M, Quail P H . Binding of phytochrome B to its nuclear signalling partner PIF3 is reversibly induced by light. Nature, 1999,400:781-784.
doi: 10.1038/23500 pmid: 10466729 |
| [14] |
Huq E . PIF4, a phytochrome-interacting bHLH factor, functions as a negative regulator of phytochrome B signaling in Arabidopsis. EMBO J, 2002,21:2441-2450.
doi: 10.1093/emboj/21.10.2441 pmid: 12006496 |
| [15] |
Kiribuchi K, Jikumaru Y, Kaku H, Minami E, Hasegawa M, Kodama O, Seto H, Okada K, Nojiri H, Yamane H . Involvement of the basic helix-loop-helix transcription factor RERJ1 in wounding and drought stress responses in rice plants. Biosci Biotechnol Biochem, 2005,69:1042-1044.
doi: 10.1271/bbb.69.1042 pmid: 15914931 |
| [16] |
Li H M, Sun J Q, Xu Y X, Jiang H L, Wu X Y, Li C Y . The bHLH-type transcription factor AtAIB positively regulates ABA response in Arabidopsis. Plant Mol Biol, 2007,65:655-665.
doi: 10.1007/s11103-007-9230-3 |
| [17] |
Jiang Y Q, Yang B, Deyholos M K . Functional characterization of the Arabidopsis bHLH92 transcription factor in abiotic stress. Mol Genet Genomics, 2009,282:503-516.
doi: 10.1007/s00438-009-0481-3 pmid: 19760256 |
| [18] |
Chinnusamy V, Ohta M, Kanrar S, Lee B H, Hong X H, Agarwal M, Zhu J K . ICE: a regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev, 2003,17:1043-1054.
doi: 10.1101/gad.1077503 pmid: 12672693 |
| [19] |
Ogo Y, Itai R N, Nakanishi H, Kobayashi T, Takahashi M, Mori S, Nishizawa N K . The rice bHLH protein OsIRO2 is an essential regulator of the genes involved in Fe uptake under Fe-deficient conditions. Plant J, 2007,51:366-377.
doi: 10.1111/j.1365-313X.2007.03149.x pmid: 17559517 |
| [20] |
Li Z X, Liu C, Zhang Y, Wang B M, Ran Q J, Zhang J R . The bHLH family member ZmPTF1 regulates drought tolerance in maize by promoting root development and abscisic acid synthesis. J Exp Bot, 2019,70:5471-5486.
doi: 10.1093/jxb/erz307 pmid: 31267122 |
| [21] |
Bailey P C, Martin C, Toledo-Otriz G, Quail P H, Huq E, Heim M A, Jakoby M, Werber M, Weisshaar B . Update on the basic helix-loop-helix transcription factor gene family in Arabidopsis thaliana. Plant Cell, 2003,15:2497-2501.
doi: 10.1105/tpc.151140 pmid: 14600211 |
| [22] |
Li X, Duan X, Jiang H X, Sun Y J, Tang Y P, Yuan Z, Guo J K, Liang W Q, Chen L, Yin J Y, Ma H, Wang J, Zhang D B . Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Physiol, 2006,141:1167-1184.
doi: 10.1104/pp.106.080580 pmid: 16896230 |
| [23] |
Zhang T T, Wei L, Zhang H S, Ma L, Li P H, Ge L, Li G . Genome-wide analysis of the basic Helix-Loop-Helix (bHLH) transcription factor family in maize. BMC Plant Biol, 2018,18:235.
doi: 10.1186/s12870-018-1441-z pmid: 30326829 |
| [24] |
Yoo S D, Cho Y H, Sheen J . Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat Protoc, 2007,2:1565-1572.
doi: 10.1038/nprot.2007.199 pmid: 17585298 |
| [25] |
Livak K J, Schmittgen T D . Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆CT method . Methods, 2001,25:402-408.
doi: 10.1006/meth.2001.1262 pmid: 11846609 |
| [26] |
Clough S J, Bent A F . Floral dip: a simplified method for Agrobacterium mediated transformation of Arabidopsis thaliana. Plant J, 1998,16:735-743.
doi: 10.1046/j.1365-313x.1998.00343.x pmid: 10069079 |
| [27] |
Chen J Q, Meng X P, Zhang Y, Xia M, Wang X P . Over-expression of OsDREB genes lead to enhanced drought tolerance in rice. Biotechnol Lett, 2008,30:2191-2198.
doi: 10.1007/s10529-008-9811-5 pmid: 18779926 |
| [28] |
Rozenn L H, Mathieu C, Dipankar C, Thomas M, Catherine B . At bHLH68 transcription factor contributes to the regulation of ABA homeostasis and drought stress tolerance in Arabidopsis. Physiol Plant, 2017,160:312-327.
doi: 10.1111/ppl.12549 pmid: 28369972 |
| [29] |
Yao P F, Li C L, Zhao X R, Li M F, Zhao H X, Guo J Y, Cai Y, Chen H, Wu Q . Overexpression of a tartary buckwheat gene, FtbHLH3, Enhances drought/oxidative stress tolerance in transgenic Arabidopsis. Front Plant Sci, 2017,8:625.
doi: 10.3389/fpls.2017.00625 pmid: 28487715 |
| [30] |
Li F, Guo S Y, Zhao Y, Chen D Z, Chong K, Xu Y Y . Overexpression of a homopeptide repeat-containing bHLH protein gene ( OrbHLH001) from Dongxiang wild rice confers freezing and salt tolerance in transgenic Arabidopsis. Plant Cell Rep, 2010,29:977-986.
doi: 10.1007/s00299-010-0883-z |
| [31] |
Zhou J, Li F, Wang J L, Ma Y, Chong K, Xu Y Y . Basic helix-loop-helix transcription factor from wild rice ( OrbHLH2) improves tolerance to salt and osmotic stress in Arabidopsis. J Plant Physiol, 2009,166:1296-1306.
doi: 10.1016/j.jplph.2009.02.007 pmid: 19324458 |
| [32] |
Liu W W, Tai H H, Li S S, Gao W, Zhao M, Xie C X, Li W X . bHLH122 is important for drought and osmotic stress resistance in Arabidopsis and in the repression of ABA catabolism. New Phytol, 2014,201:1192-1204.
doi: 10.1111/nph.12607 |
| [33] |
Xu W R, Zhang N B, Jiao Y T, Li R M, Xiao D M, Wang Z . The grapevine basic helix-loop-helix (bHLH) transcription factor positively modulates CBF-pathway and confers tolerance to cold-stress in Arabidopsis. Mol Biol Rep, 2014,41:5329-5342.
doi: 10.1007/s11033-014-3404-2 |
| [34] |
Zhao Q, Xiang X, Liu D, Yang A, Wang Y . Tobacco transcription factor NtbHLH123 confers tolerance to cold stress by regulating the NtCBF pathway and reactive oxygen species homeostasis. Front Plant Sci, 2018,9:381.
doi: 10.3389/fpls.2018.00381 pmid: 29643858 |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [4] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [5] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [6] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [7] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [8] | 朱峥, 王田幸子, 陈悦, 刘玉晴, 燕高伟, 徐珊, 马金姣, 窦世娟, 李莉云, 刘国振. 水稻转录因子WRKY68在Xa21介导的抗白叶枯病反应中发挥正调控作用[J]. 作物学报, 2022, 48(5): 1129-1140. |
| [9] | 陈悦, 孙明哲, 贾博为, 冷月, 孙晓丽. 水稻AP2/ERF转录因子参与逆境胁迫应答的分子机制研究进展[J]. 作物学报, 2022, 48(4): 781-790. |
| [10] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [11] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [12] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [13] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [14] | 黄成, 梁晓梅, 戴成, 文静, 易斌, 涂金星, 沈金雄, 傅廷栋, 马朝芝. 甘蓝型油菜BnAPs基因家族成员全基因组鉴定及分析[J]. 作物学报, 2022, 48(3): 597-607. |
| [15] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
|
||