欢迎访问作物学报,今天是
作物学报 ›› 2020, Vol. 46 ›› Issue (9): 1359-1367.doi: 10.3724/SP.J.1006.2020.03005
石慧敏1( ), 蒋成功1,2, 王红武1, 马庆2, 李坤1, 刘志芳1, 吴宇锦1, 李树强1, 胡小娇1,*(), 黄长玲1,*()
), 蒋成功1,2, 王红武1, 马庆2, 李坤1, 刘志芳1, 吴宇锦1, 李树强1, 胡小娇1,*(), 黄长玲1,*()
SHI Hui-Min1(), JIANG Cheng-Gong1,2, WANG Hong-Wu1, MA Qing2, LI Kun1, LIU Zhi-Fang1, WU Yu-Jin1, LI Shu-Qiang1, HU Xiao-Jiao1,*(), HUANG Chang-Ling1,*()
摘要:
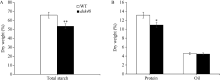
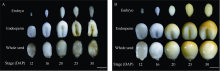
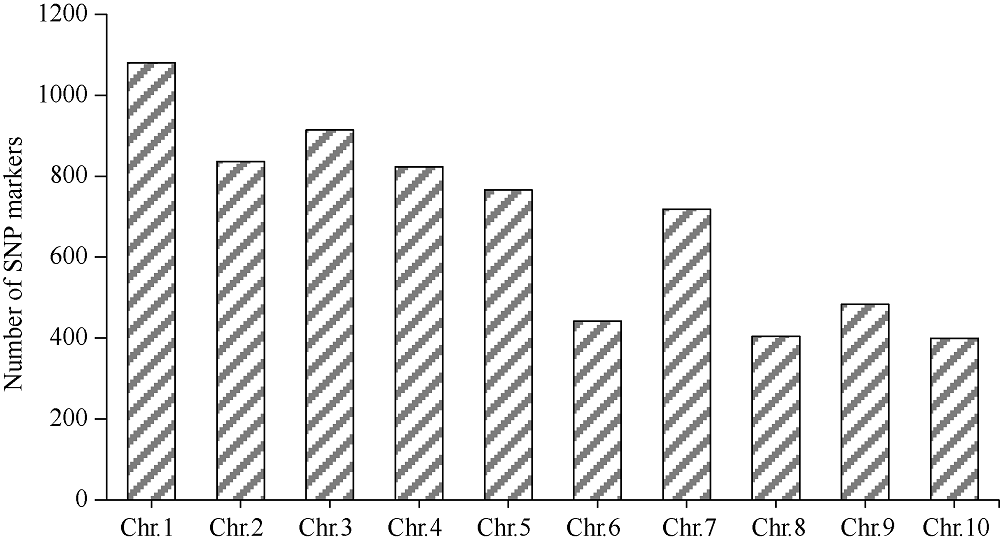
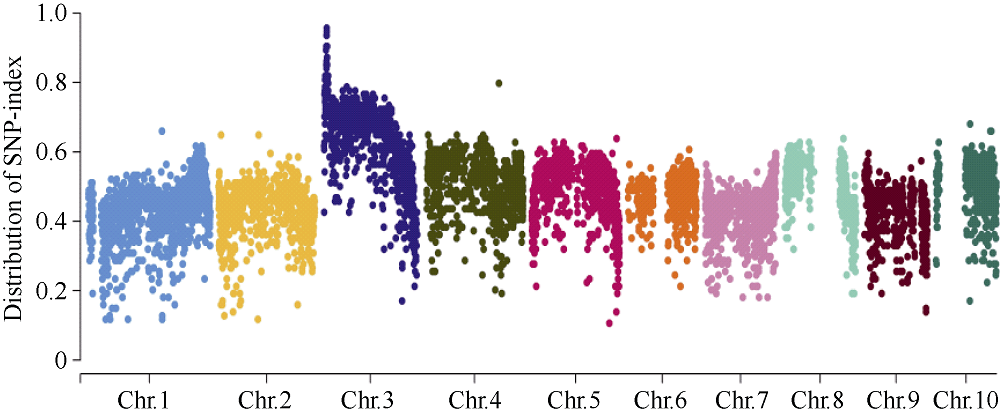
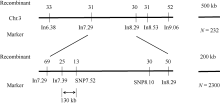
籽粒是玉米的主要营养储存器官, 也是禾本科植物种子发育研究的模式器官。本研究对玉米自交系郑58进行甲基磺酸乙酯(EMS)诱变, 获得一个稳定遗传的籽粒缺陷突变体, 命名为defective kernel 48 (dek48)。该突变体籽粒皱缩扁小, 百粒重显著降低, 胚和胚乳发育严重缺陷, 不能成苗。在玉米授粉后12 d即可观察到明显的发育缺陷, 表明该突变发生在籽粒发育的早期阶段。扫描电镜观察发现dek48与野生型相比淀粉粒显著变小。石蜡切片显微观察发现dek48淀粉胚乳填充不饱满, 糊粉层细胞发育不规则。遗传学分析表明, 该突变性状受隐性单基因控制。进一步构建F2遗传定位群体, 将该突变体基因精细定位于3号染色体7.39 Mb~7.52 Mb之间。生物信息学分析发现该区间内有6个开放阅读框, 暂未发现与籽粒发育有关的已知基因, 后续将通过测序和基因表达分析进一步确定候选基因。
| [1] | 李穆, 孟令聪, 郑淑波, 周德龙, 刘宏伟, 王敏, 何欢, 路明. “十二五”以来我国玉米分子育种研究进展. 玉米科学, 2019,27(6):1-6. |
| Li M, Meng L C, Zeng S B, Zhou D L, Liu H W, Wang M, He H, Lu M. Advances in molecular breeding of maize in China since the 12th Five-Year Plan. J Maize Sci, 2019,27(6):1-6 (in Chinese with English abstract). | |
| [2] |
Neuffer M G, Sheridan W F. Defective kernel mutants of maize: I. Genetic and lethality studies. Genetics, 1980,95:929-944.
pmid: 17249053 |
| [3] |
Janice K C, William F S. Characterization of the two maize embryo-lethal defective kernel mutants rgh*-1210 and fl*-1253b: effects on embryo and gametophyte development. Genetics, 1988,120:279-90.
pmid: 17246478 |
| [4] |
Sheridan W F, Neuffer M G. Defective kernel mutants of maize: II. Morphological and embryo culture studies. Genetics, 1980,95:945-60.
pmid: 17249054 |
| [5] | Scanlon M J, Stinard P S, James M G, Myers A M, Robertson D S. Genetic analysis of 63 mutations affecting maize kernel development isolated from mutator stocks. Genetics, 1994,136:281-94. |
| [6] |
Lurin C, Andrés C, Aubourg S, Bellaoui M, Bitton F, Bruyère C, Caboche M, Debast C, Gualberto J, Hoffmann B, Lecharny A, LeRet M, MartinMagniette M L, Mireau H, Peeters N, Renou J P, Szurek B, Taconnat L, Small I. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell, 2004,16:2089-2103.
pmid: 15269332 |
| [7] | Colcombet J, Lopez-Obando M, Heurtevin L, Bernard C, Martin K, Berthomé R, Lurin C. Systematic study of subcellular localization of Arabidopsis PPR proteins confirms a massive targeting to organelles. RNA Biol, 2013,10:1557-1575. |
| [8] |
Nikolay M, Virginie G, Jörg M, Udo W, Reinhold B. An essential pentatricopeptide repeat protein facilitates 59 maturation and translation initiation of rps3 mRNA in maize mitochondria. Plant Cell, 2012,24:3087-3105.
pmid: 22773745 |
| [9] |
Qi W, Yang Y, Feng X, Zhang M, Song R. Mitochondrial function and maize kernel development requires dek2, a pentatricopeptide repeat protein involved in nad1 mRNA splicing. Genetics, 2017,205:239-249.
doi: 10.1534/genetics.116.196105 pmid: 27815362 |
| [10] |
Qi W, Tian Z, Lu L, Chen X, Chen X, Zhang W, Song R. Editing of mitochondrial transcripts nad3 and cox2 by dek10 is essential for mitochondrial function and maize plant development. Genetics, 2017,205:1489-1501.
doi: 10.1534/genetics.116.199331 pmid: 28213476 |
| [11] | Chen X, Feng F, Qi W, Xu L, Yao D, Wang Q, Song R. dek35 encodes a PPR protein that affects cis-splicing of mitochondrial nad4 intron 1 and seed development in maize. Mol Plant, 2017,10:427-441. |
| [12] |
Wang G, Zhong M, Shuai B, Song J, Zhang J, Han L, Ling H, Tang Y, Wang G, Song R. E+ subgroup PPR protein defective kernel 36 is required for multiple mitochondrial transcripts editing and seed development in maize and Arabidopsis. New Phytol, 2017,214:1563-1578.
doi: 10.1111/nph.14507 pmid: 28277611 |
| [13] |
Dai D, Luan S, Chen X, Wang Q, Feng Y, Zhu C, Qi W, Song R. Maize dek37 encodes a P-type PPR protein that affects cis-splicing of mitochondrial nad2 intron 1 and seed developmen. Genetics, 2018,208:1069-1082.
pmid: 29301905 |
| [14] |
Li X, Gu W, Sun S, Chen Z, Chen J, Song W, Zhao H, Lai J. Defective Kernel 39 encodes a PPR protein required for seed development in maize. J Integr Plant Biol, 2018,60:45-64.
doi: 10.1111/jipb.12602 pmid: 28981206 |
| [15] | Ren R C, Lu X, Zhao Y J, Wei Y M, Wang L L, Zhang L, Zhang W T, Zhang C, Zhang X S, Zhao X Y. Pentatricopeptide repeat protein dek40 is required for mitochondrial function and kernel development in maize. J Exp Bot, 2019,70:6163-6379. |
| [16] | Zhu C, Jin G, Fang P, Zhang Y, Feng X, Tang Y, Qi W, Song R. Maize pentatricopeptide repeat protein dek41 affects cis-splicing of mitochondrial nad4 intron 3 and is required for normal seed development. J Exp Bot, 2019,70:3795-3808. |
| [17] |
He Y H, Wang J G, Qi W W, Song R T. Maize dek15 encodes the cohesion-loading complex subunit SCC4 and is essential for chromosome segregation and kernel development. Plant Cell, 2019,31:465-485.
doi: 10.1105/tpc.18.00921 pmid: 30705131 |
| [18] | Dai D W, Tong H Y, Cheng L J, Peng F, Zhang T T, Qi W W, Song R T. Maize dek33 encodes a pyrimidine reductase in riboflavin biosynthesis that is essential for oil-body formation and ABA biosynthesis during seed development. J Exp Bot, 2019,19:5173-5187. |
| [19] |
Nelson G, Li Y B, Hugo K D, Joachim M. Maize defective kernel mutant generated by insertion of a Ds element in a gene encoding a highly conserved TTI2 cochaperone. Proc Natl Acad Sci USA, 2017,114:5165-5170.
doi: 10.1073/pnas.1703498114 pmid: 28461460 |
| [20] |
Zuo Y, Feng F, Qi W W, Song R T. dek42 encodes an RNA-binding protein that affects alternative pre-mRNA splicing and maize kernel development. J Integr Plant Biol, 2019,61:728-748.
doi: 10.1111/jipb.12798 pmid: 30839161 |
| [21] |
Qi W W, Lu L, Huang S C, Song R T. Maize dek44 encodes mitochondrial ribosomal protein L9 and is required for seed development. Plant Physiol, 2019,180:2106-2119.
doi: 10.1104/pp.19.00546 pmid: 31182559 |
| [22] | 李艺, 铁双贵, 朱卫红, 齐建双, 卢彩霞, 孙建军, 周柯. 快速CTAB法提取玉米种子胚基因组DNA. 河南农业科学, 2008,37(2):17-20. |
| Li Y, Tie S G, Zhu W H, Qi J S, Lu C X, Sun J J, Zhou K. DNA extraction from Maize embryo by a new rapid CTAB method. Henan Acad Agric Sci, 2008,37(2):17-20 (in Chinese with English abstract). | |
| [23] | Guo Z F, Wang H W, Tao J J, Ren Y H, Xu C, Wu K S, Zou C, Zhang J N, Xu Y B. Development of multiple SNP marker panels affordable to breeders through genotyping by target sequencing (GBTS) in maize. Mol Breed, 2019,39:37. |
| [24] |
Abe A, Kosugi S, Yoshida K, Natsume S, Takagi H, Kanzaki H, Matsumura H, Yoshida K, Mitsuoka C, Tamiru M, Innan H, Cano L, Kamoun S, Terauchi R. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat Biotechnol, 2012,30:174-178.
pmid: 22267009 |
| [25] | 高玉峰, 张攀, 郝晓敏, 严建兵, 李健生, 杨小红. 一种快速提取玉米大群体基因组DNA的方法. 中国农业大学学报, 2011,16(6):32-36. |
| Gao Y F, Zhang P, Hao X M, Yan J B, Li J S, Yang X H. A rapid DNA extiaction method for large maize populations. J China Agric Univ, 2011,16(6):32-36 (in Chinese with English abstract). | |
| [26] |
Birchler J A. Dosage analysis of maize endosperm development. Annu Rev Genet, 1993,27:181-204.
doi: 10.1146/annurev.ge.27.120193.001145 pmid: 8122901 |
| [27] |
Hou H L, Wang P, Zhang H, Wen H, Gao F, Ma N J, Wang Q, Li L J. Histone acetylation is involved in gibberellin-regulated s odCp gene expression in maize aleurone layers. Plant Cell Physiol, 2015,56:2139-2149.
doi: 10.1093/pcp/pcv126 pmid: 26374791 |
| [28] |
Yi G, Lauter A M, Scott M P, Becraft P W. The thick aleurone1 mutant defines a negative regulation of maize aleurone cell fate that functions downstream of defective kernel1. Plant Physiol, 2011,156:1826-36.
doi: 10.1104/pp.111.177725 pmid: 21617032 |
| [29] |
Tian Q, Olsen L, Sun B, Lid S E, Brown R C, Lemmon B E, Fosnes K, Gruis D F, Opsahl H G, Otegui M S, Olsen O A. Subcellular localization and functional domain studies of DEFECTIVE KERNEL1 in maize and Arabidopsis suggest a model for aleurone cell fate specification involving CRINKLY4 and SUPERNUMERARY ALEURONE LAYER1. Plant Cell, 2007,19:3127-45.
doi: 10.1105/tpc.106.048868 pmid: 17933905 |
| [30] |
Becraft P W, Stinard P S, McCarty D R. CRINKLY4: A TNFR-like receptor kinase involved in maize epidermal differentiation. Science, 1996,273:1406-1409.
doi: 10.1126/science.273.5280.1406 pmid: 8703079 |
| [31] |
Shen B, Li C J, Min Z, Meeley R B, Tarczynski M C, Olsen O A. sal1 determines the number of aleurone cell layers in maize endosperm and encodes a class E vacuolar sorting protein. Proc Natl Acad Sci USA, 2003,100:6552-6557.
doi: 10.1073/pnas.0732023100 pmid: 12750475 |
| [32] |
Yi G, Lauter A M, Scott M P, Becraft P W. The thick aleurone1 mutant defines a negative regulation of maize aleurone cell fate that functions downstream of defective kernel1. Plant Physiol, 2011,156:1826-1836.
doi: 10.1104/pp.111.177725 pmid: 21617032 |
| [33] |
Guo D, Zhang J, Wang X, Han X, Wei B, Wang J, Li B, Yu H, Huang Q, Gu H, Qu L J, Qin G. The WRKY transcription factor WRKY71/EXB1 controls shoot branching by transcriptionally regulating RAX genes in Arabidopsis. Plant Cell, 2015,27:3112-3127.
doi: 10.1105/tpc.15.00829 pmid: 26578700 |
| [34] |
Sarris P F, Duxbury Z, Huh S U, Ma Y, Segonzac C, Sklenar J, Derbyshire P, Cevik V, Rallapalli G, Saucet S B, Wirthmueller L, Menke F L H, Sohn K H, Jones J D G. A plant immune receptor detects pathogen effectors that target WRKY transcription factors. Cell, 2015,161:1089-1100.
doi: 10.1016/j.cell.2015.04.024 pmid: 26000484 |
| [35] |
Chen L, Yang Y, Liu C, Zheng Y, Xu M, Wu N, Sheng J, Shen L. Characterization of WRKY transcription factors in Solanum lycopersicum reveals collinearity and their expression patterns under cold treatment. Biochem Biophys Res Commun, 2015,464:962-968.
doi: 10.1016/j.bbrc.2015.07.085 pmid: 26196744 |
| [36] | 李琪, 李烨, 牛芳芳, 郭小华, 赵新杰, 吴相民, 杨博, 江元清. 拟南芥转录因子基因WRKY72的特性分析及其抗逆功能鉴定. 农业生物技术学报, 2019,27:191-203. |
| Li Q, Li Y, Niu F F, Guo X H, Zhao X J, Wu X M, Yang B, Jiang Y Q. Characterization and stress-resistance functional identification of transcription factor gene WRKY72 in Arabidopsis thaliana. J Agric Biotechnol, 2019,27:191-203 (in Chinese with English abstract). | |
| [37] |
Weits D A, Giuntoli B, Kosmacz M, Parlanti S, Hubberten H M, Riegler H, Hoefgen R, Perata P, van Dongen J T, Licausi F. Plant cysteine oxidases control the oxygen-dependent branch of the N-end-rule pathway. Nat Commun, 2014,5:3425.
doi: 10.1038/ncomms4425 pmid: 24599061 |
| [38] | 秘彩莉, 刘旭, 张学勇. F-box蛋白在植物生长发育中的功能. 遗传, 2006,28:1337-1342. |
| Mi C I, Liu X, Zhang X Y. The function of F-box protein in plant growth and development. Hereditas, 2006,28:1337-1342 (in Chinese with English abstract). | |
| [39] |
Mitchell R A, Dupree P, Shewry P R. A novel bioinformatics approach identifies candidate genes for the synthesis and feruloylation of arabinoxylan. Plant Physiol, 2007, 144:43-53.
doi: 10.1104/pp.106.094995 pmid: 17351055 |
| [40] |
Molinari H B C, Pellny T K, Freeman J, Shewry P R, Mitchell, R A C. Grass cell wall feruloylation: distribution of bound ferulate and candidate gene expression in Brachypodium distachyon. Front Plant Sci, 2013,4:50.
doi: 10.3389/fpls.2013.00050 pmid: 23508643 |
| [41] |
Dascher C, Matteson J, Walch W E. Syntaxin5 regulates endoplasmatic reticulum to Golgi transport. J Biol Chem, 1994,269:29363-29366.
pmid: 7961911 |
| [42] |
Morita M T, Tasaka M. Gravity sensing and signaling. Curr Opin Plant Biol, 2004,7:712-718.
doi: 10.1016/j.pbi.2004.09.001 pmid: 15491921 |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 郑崇珂, 周冠华, 牛淑琳, 和亚男, 孙伟, 谢先芝. 水稻早衰突变体esl-H5的表型鉴定与基因定位[J]. 作物学报, 2022, 48(6): 1389-1400. |
| [4] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [5] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [6] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [7] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [8] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [9] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [10] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [11] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [12] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [13] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [14] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [15] | 张倩, 韩本高, 张博, 盛开, 李岚涛, 王宜伦. 控失尿素减施及不同配比对夏玉米产量及氮肥效率的影响[J]. 作物学报, 2022, 48(1): 180-192. |
|
||