欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (8): 1905-1913.doi: 10.3724/SP.J.1006.2022.11069
杜启迪1,2( ), 郭会君2, 熊宏春2, 谢永盾2, 赵林姝2, 古佳玉2, 赵世荣2, 丁玉萍2, 宋希云1, 刘录祥2,*()
), 郭会君2, 熊宏春2, 谢永盾2, 赵林姝2, 古佳玉2, 赵世荣2, 丁玉萍2, 宋希云1, 刘录祥2,*()
DU Qi-Di1,2(), GUO Hui-Jun2, XIONG Hong-Chun2, XIE Yong-Dun2, ZHAO Lin-Shu2, GU Jia-Yu2, ZHAO Shi-Rong2, DING Yu-Ping2, SONG Xi-Yun1, LIU Lu-Xiang2,*()
摘要:
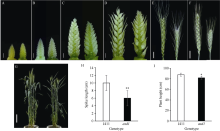
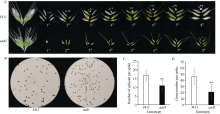
穗粒数是小麦产量三要素建成的关键因子, 深入挖掘穗部发育调控基因有助于培育高产小麦品种。以小麦品种京411为野生型, 经EMS诱变获得了表型稳定的小穗退化突变体asd1 (apical spikelet degeneration 1)。该突变体表现顶端小穗明显退化, 穗长缩短了约40%, 结实小穗数减少了约35%, 穗粒数显著减少了54%, 同时株高也明显降低。利用京411×asd1遗传群体的F2和F3代表型数据分析表明, 顶端小穗退化性状受1对主效隐性基因控制。采用混合群体分离分析法(BSA), 结合测序所得SNP位点, 在7A染色体上开发了7个KASP标记, 将目标突变基因定位在7A染色体短臂9.91 Mb物理区间内, 遗传距离为17.62 cM, 推断该区段存在一个新的控制小麦花器官发育及穗部形态发育的重要基因。本研究所鉴定的小麦穗发育控制区段有助于深入解析小麦小穗形成的遗传基础, 为进一步揭示小麦产量形成的分子机理提供突变基因。
| [1] | 余泽高, 许立俊. 小麦穗部性状间的相关及穗粒数改良途径的研究. 湖北农业科学, 2002, (6): 38-40. |
| Yu Z G, Xu L J. Analysis of correlation of spike-section characteristics and way of reform kernel number in wheat. Hubei Agric Sci, 2002, (6): 38-40. | |
| [2] | 王兆龙, 曹卫星, 戴廷波. 小麦穗粒数形成的基因型差异及增粒途径分析. 作物学报, 2001, 27: 236-242. |
| Wang Z L, Cao W X, Dai T B. Genotypic differences in formation of kernel number per spike and analysis of improvement approaches in wheat. Acta Agron Sin, 2001, 27: 236-242. | |
| [3] |
Zhang B, Liu X, Xu W N, Chang J Z, Li A, Mao X G, Zhang X Y, Jing R L. Novel function of a putative MOC1 ortholog associated with spikelet number per spike in common wheat. Sci Rep, 2015, 5: 12211.
doi: 10.1038/srep12211 pmid: 26197925 |
| [4] | 徐伟娜. 小麦穗发育相关基因TaSPL20的生物学功能分析. 中国农业科学院硕士学位论文,北京, 2017. |
| Xu W N. Biological Function of Ear Development Related Gene TaSPL20 from Wheat (Triticum aestivum L.). MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2017. | |
| [5] |
Zhang B, Xu W N, Liu X, Mao X G, Li A, Wang J Y, Chang X P, Zhang X Y, Jing R L. Functional conservation and divergence among homoeologs of TaSPL20 and TaSPL21, two SBP box genes governing yield-related traits in hexaploid wheat. Plant Physiol, 2017, 174: 1177-1191.
doi: 10.1104/pp.17.00113 pmid: 28424214 |
| [6] |
Wang Y G, Yu H P, Tian C H, Sajjad M, Gao C X, Tong Y P, Wang X F, Jiao Y L. Transcriptome association identifies regulators of wheat spike architecture. Plant Physiol, 2017, 175: 746-757.
doi: 10.1104/pp.17.00694 |
| [7] |
Oxana D, Caroline P, Richard S, Petr M, Ekaterina B, Florent M, Audrey C, Nobuyoshi W, Elisa P, Nadine G, Véronique G, Charles P, Yuriy L O, Alexander A K, Hélène B, Elena S, Lyudmila L, Jerome S. FRIZZY PANICLE drives supernumerary spikelets in bread wheat. Plant Physiol, 2015, 167: 189-199.
doi: 10.1104/pp.114.250043 |
| [8] |
Li Y P, Li L, Zhao M C, Guo L, Guo X X, Zhao D, Batool A, Dong B D, Xu H X, Cui S J, Zhang A M, Fu X D, Li J M, Jing R L, Liu X G. Wheat FRIZZY PANICLE activates VERNALIZATION1-A and HOMEOBOX4-A to regulate spike development in wheat. Plant Biotechnol J, 2021, 19: 1141-1154.
doi: 10.1111/pbi.13535 |
| [9] |
Dixon L E, Greenwood J R, Bencivenga S, Zhang P, Cockram J, Mellers G, Ramm K, Cavanagh C, Swain S M, Boden S A. TEOSINTE BRANCHED1 regulates inflorescence architecture and development in bread wheat (Triticum aestivum). Plant Cell, 2018, 30: 563-581.
doi: 10.1105/tpc.17.00961 |
| [10] |
Boden S A, Cavanagh C, Cullis B R, Ramm K, Greenwood J, Finnegan E J, Trevaskis B, Swain S M. Ppd-1 is a key regulator of inflorescence architecture and paired spikelet development in wheat. Nat Plant, 2015, 1: 14016.
doi: 10.1038/nplants.2014.16 |
| [11] |
Okada T, Jayasinghe J E A R M, Eckermann P, Watson-Haigh N S, Warner P, Hendrikse Y, Baes M, Tucker E J, Laga H, Kato K, Albertsen M, Wolters P, Fleury D, Baumann U, Whitford R. Effects of Rht-B1and Ppd-D1 loci on pollinator traits in wheat. Theor Appl Genet, 2019, 132: 1965-1979.
doi: 10.1007/s00122-019-03329-w |
| [12] | 周丽敏. 小麦穗发育异常相关基因 TaSDA1的定位研究. 西北农林科技大学硕士学位论文,陕西杨凌, 2016. |
| Zhou L M. Mapping Research of TASDA1 Gene of Triricum aestivum Spike Development Atrophy1. MS Thesis of Northwest A&F University of Agriculture and Forestry, Yangling, Shaanxi, China, 2016. | |
| [13] | 蒋方山, 郭营, 许云峰, 李瑞军, 李斯深. EMS诱变的小麦基部小穗不孕突变体的鉴定与小穗形态发育. 麦类作物学报, 2008, 28: 249-253. |
| Jiang F S, Guo Y, Xu Y F, Li R J, Li S S. Identification of a distal spikelet sterility mutant in wheat and spikelet morphological development. J Triticeae Crops, 2008, 28: 249-253. | |
| [14] | 顾晶晶. 小麦穗发育突变体SMS1的鉴定和基因定位. 河南农业大学硕士学位论文,河南郑州, 2017. |
| Gu J J. Identification and Genetic Mapping of a Sterile and Malformed Spike 1 Gene in Common Wheat. MS Thesis of Henan Agricultural University, Zhengzhou, Henan, China, 2017. | |
| [15] |
Sakuma S, Golan G, Guo Z F, Ogawa T, Tagiri A, Sugimoto K, Bernhardt N, Brassac J, Mascher M, Hensel G, Ohnishi S, Jinno H, Yamashita Y, Ayalon I, Peleg Z, Schnurbusch T, Komatsuda T. Unleashing floret fertility in wheat through the mutation of a homeobox gene. Proc Natl Acad Sci USA, 2019, 116: 5182-5187.
doi: 10.1073/pnas.1815465116 |
| [16] |
Xia C, Zhang L C, Zou C, Gu Y Q, Duan J L, Zhao G Y, Wu J J, Liu Y, Fang X H, Gao L F, Jiao Y N, Sun J Q, Pan Y H, Liu X, Jia J Z, Kong X Y. A TRIM insertion in the promoter of Ms2 causes male sterility in wheat. Nat Commun, 2017, 8:15407.
doi: 10.1038/ncomms15407 |
| [17] |
Ni F, Qi J, Hao Q Q, Bo L, Luo M C, Wang Y, Chen F J, Wang S Y, Zhang C Z, Epstein L, Zhao X Y, Wang H G, Zhang X S, Chen C X, Sun L Z, Fu D L. Wheat Ms2encodes for an orphan protein that confers male sterility in grass species. Nat Commun, 2017, 8: 15121.
doi: 10.1038/ncomms15121 |
| [18] | 翟虎渠, 刘秉华. 矮败小麦创制与应用. 中国农业科学, 2009, 42: 4127-4131. |
| Zhai H Q, Liu B H. The innovation of dwarf male sterile wheat and its application in wheat breading. Sci Agric Sin, 2009, 42: 4127-4131. | |
| [19] |
Yan L L, Loukoianov A, Tranquilli G, Helguera M, Fahima T, Dubcovsky J. Positional cloning of the wheat vernalization gene VRN1. Proc Natl Acad Sci USA, 2003, 100: 6263-6268.
doi: 10.1073/pnas.0937399100 |
| [20] |
Li C X, Lin H Q, Chen A, Lau M, Jernstedt J, Dubcovsky J. Wheat VRN1, FUL2 and FUL3play critical and redundant roles in spikelet development and spike determinacy. Development, 2019, 146: dev175398.
doi: 10.1242/dev.175398 |
| [21] |
Jill C P, Elizabeth A K. Reconstructing the evolutionary history of paralogous APETALA1/FRUITFULL-like genes in grasses (Poaceae). Genetics, 2006, 174: 421-437.
doi: 10.1534/genetics.106.057125 |
| [22] | Sun C F, Niu Y C, Ye X, Dong J J, Hu W S, Zeng Q K, Chen Z H, Tian Y Y, Zhang J, Lu M X. Development of a high-density linkage map and mapping of the three-pistil gene (Pis1) in wheat using GBS markers. BMC Gnomics, 2017, 18: 567. |
| [23] |
Zou C, Wang P X, Xu Y B. Bulked sample analysis in genetics, genomics and crop improvement. Plant Biotechnol J, 2016, 14: 1941-1955.
doi: 10.1111/pbi.12559 |
| [24] |
Robert K, Nicholas B, Ricardo R G, Jane A C, Archana P, Keywan H P, Cristobal U, Andrew L P. Mutation scanning in wheat by exon capture and next-generation sequencing. PLoS One, 2015, 10: e0137549.
doi: 10.1371/journal.pone.0137549 |
| [25] |
Yao Z, You F M, N' Diaye A, Knox R E, McCartney C, Hiebert C W, Pozniak C, Xu W. Evaluation of variant calling tools for large plant genome re-sequencing. BMC Bioinformatics, 2020, 21: 360.
doi: 10.1186/s12859-020-03704-1 |
| [26] |
Hill J T, Demarest B L, Bisgrove B W, Bushra G, Su Y C, Yost H J. MMAPPR: mutation mapping analysis pipeline for pooled RNA-seq. Genome Res, 2013, 23: 687-697.
doi: 10.1101/gr.146936.112 |
| [27] | 张顺麟. 冬小麦淀粉合成关键基因TaSSIVb特性分析与等位变异挖掘. 中国农业科学院硕士学位论文,北京, 2019. |
| Zhang S L. Characterization of Starch Synthesis Key Gene TaSSIVb and Mining of its Mutation Alleles in Winter Wheat. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2019. | |
| [28] |
Meng L, Li H H, Zhang L Y, Wang J K. QTL IciMapping: integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J, 2015, 3: 269-283.
doi: 10.1016/j.cj.2015.01.001 |
| [29] |
Koji M, Shigeo T, Hironori K, Yasunari O. Pistillody, homeotic transformation of stamens into pistil-like structures, caused by nuclear-cytoplasm interaction in wheat. Plant J, 2002, 29: 169-181.
doi: 10.1046/j.0960-7412.2001.01203.x |
| [30] | 李存东, 曹卫星, 戴廷波, 严美春, 王兆龙. 小麦小花原基分化和退化的动态模式与特征. 中国农业科学, 1999, 32(5): 98-100. |
| Li C D, Cao W X, Dai T B, Yan M L, Wang Z L. Study on dynamic models and characteristics of floret primordium differentiation and degeneration in wheat. Sci Agric Sin, 1999, 32(5): 98-100. | |
| [31] |
Kuzay S, Xu Y F, Zhang J L, Katz A, Pearce S, Su Z Q, Fraser M, Anderson J A, Brown-Guedira G, Witt N D, Haugrud A P, Faris J D, Akhunov E, Bai G H, Dubcovsky J. Identification of a candidate gene for a QTL for spikelet number per spike on wheat chromosome arm 7AL by high-resolution genetic mapping. Theor Appl Genet, 2019, 132: 2689-2705.
doi: 10.1007/s00122-019-03382-5 |
| [32] |
Corsi B, Obinu L, Zanella C M, Cutrupi S, Day R, Geyer M, Lillemo M, Lin M, Mazza L, Percival-Alwyn L, Stadlmeier M, Mohler V, Hart L, Cockram J. Identification of eight QTL controlling multiple yield components in a German multi-parental wheat population, including Rht24, WAPO-A1, WAPO-B1 and genetic loci on chromosomes 5A and 6A. Theor Appl Genet, 2021, 134: 1435-1454.
doi: 10.1007/s00122-021-03781-7 |
| [33] | Tang Y L, Li J, Wu Y Q, Wei H T, Li C S, Yang W Y, Chen F. Identification of QTLs for yield-related traits in the recombinant inbred line population derived from the cross between a synthetic hexaploid wheat-derived variety Chuanmai 42 and a Chinese elite variety Chuannong 16. J Integr Agric, 2011, 10: 1665-1680. |
| [34] |
Jantasuriyarat C, Vales M I, C Watson J W, Riera-Lizarazu O. Identification and mapping of genetic loci affecting the free-threshing habit and spike compactness in wheat (Triticum aestivum L.). Theor Appl Genet, 2004, 108: 261-273.
pmid: 13679977 |
| [35] | 宋全昊, 刘路平, 李法计, 田芳慧, 孙道杰. 小麦穗部发育多效基因的遗传分析与基因定位. 西北植物学报, 2013, 33: 643-648. |
| Song Q H, Liu L P, Li F J, Tian F H, Sun D J. Genetic analysis and gene mapping of the spike development pleiotropic gene in wheat. Acta Bot Boreali-Occident Sin, 2013, 33: 643-648. | |
| [36] |
Tomio T, Kenji N, Kazuko M, Tatsuro H. A gene controlling the number of primary rachis branches also controls the vascular bundle formation and hence is responsible to increase the harvest index and grain yield in rice. Theor Appl Genet, 2010, 120: 875-893.
doi: 10.1007/s00122-009-1218-8 pmid: 20151298 |
| [37] |
Ikeda-Kawakatsu K, Maekawa M, Izawa T, Itoh J I, Nagato Y. ABERRANT PANICLE ORGANIZATION 2/RFL, the rice ortholog of Arabidopsis LEAFY, suppresses the transition from inflorescence meristem to floral meristem through interaction with APO1. Plant J, 2012, 69: 168-180.
doi: 10.1111/j.1365-313X.2011.04781.x |
| [38] |
Huang L J, Hua K, Xu R, Zeng D L, Wang R C, Dong G J, Zhang G Z, Lu X L, Fang N, Wang D K, Duan P G, Zhang B L, Liu Z P, Li N, Luo Y H, Qian Q, Yao S G, Li Y H. The LARGE2-APO1/APO2 regulatory module controls panicle size and grain number in rice. Plant Cell, 2021, 33: 1212-1228.
doi: 10.1093/plcell/koab041 |
| [39] |
Akiko Y, Yoshihiro O, Hidemi K, Fumio T S, Hiro Y H. ABERRANT SPIKELET and PANICLE1, encoding a TOPLESS- related transcriptional corepressor, is involved in the regulation of meristem fate in rice. Plant J, 2021, 70: 327-339.
doi: 10.1111/j.1365-313X.2011.04872.x |
| [40] |
Heng Y Q, Wu C Y, Long Y, Luo S, Ma J, Chen J, Liu J F, Zhang H, Ren Y L, Wang M, Tan J J, Zhu S S, Wang J L, Lei C L, Zhang X, Guo X P, Wang H Y, Cheng Z J, Wan J M. OsALMT7 maintains panicle size and grain yield in rice by mediating malate transport. Plant Cell, 2018, 30: 889-906.
doi: 10.1105/tpc.17.00998 |
| [41] |
Wang Q L, Sun A Z, Chen S T, Chen L S, Guo F Q. SPL6 represses signaling outputs of ER stress in control of panicle cell death in rice. Nat Plant, 2018, 4: 280-288.
doi: 10.1038/s41477-018-0131-z |
| [42] |
Zafar S A, Patil S B, Uzair M, Fang J J, Zhao J F, Guo T T, Yuan S J, Uzair M, Luo Q, Shi J X, Schreiber L, Li X Y. DEGENERATED PANICLE AND PARTIAL STERILITY 1 (DPS1) encodes a cystathionine β-synthase domain containing protein required for anther cuticle and panicle development in rice. New Phytol, 2020, 225: 356-375.
doi: 10.1111/nph.16133 |
| [43] |
Jose F G, Behzad T, Zoe A W. Anther and pollen development: a conserved developmental pathway. J Integr Plant Biol, 2015, 57: 876-891.
doi: 10.1111/jipb.12425 |
| [44] |
Liu Z, Lin S, Shi J X, Yu J, Zhu L, Yang X J, Zhang D B, Liang W Q. Rice No Pollen 1 (NP1) is required for anther cuticle formation and pollen exine patterning. Plant J, 2017, 91: 263-277.
doi: 10.1111/tpj.13561 |
| [1] | 张一铎, 李国强, 孔忠新, 王玉泉, 李小利, 茹振钢, 贾海燕, 马正强. 基因聚合选育抗赤霉病小麦新品系百农4299[J]. 作物学报, 2022, 48(9): 2221-2227. |
| [2] | 谭照国, 苑少华, 李艳梅, 白建芳, 岳洁茹, 刘子涵, 张天豹, 赵福永, 赵昌平, 许本波, 张胜全, 庞斌双, 张立平. 小麦TaPIP1基因的克隆及其在花药开裂中的潜在功能分析[J]. 作物学报, 2022, 48(9): 2242-2254. |
| [3] | 冯子恒, 李晓, 段剑钊, 高飞, 贺利, 杨天聪, 戎亚思, 宋莉, 尹飞, 冯伟. 基于特征波段选择和机器学习的小麦白粉病高光谱遥感监测[J]. 作物学报, 2022, 48(9): 2300-2314. |
| [4] | 曹际玲, 曾青, 朱建国. 不同品种小麦灌浆期旗叶光合特性及光合基因表达对臭氧浓度升高的响应[J]. 作物学报, 2022, 48(9): 2339-2350. |
| [5] | 李永波, 崔德周, 黄琛, 隋新霞, 樊庆琦, 楚秀生. 高度特异性小麦ATG8抗体的研制及其在细胞自噬检测中的应用[J]. 作物学报, 2022, 48(9): 2390-2399. |
| [6] | 王云奇, 高福莉, 李傲, 郭同济, 戚留冉, 曾寰宇, 赵建云, 王笑鸽, 高国英, 杨佳鹏, 白金泽, 马亚欢, 梁月馨, 张睿. 小麦花后穗部温度变化规律及其与产量的关系[J]. 作物学报, 2022, 48(9): 2400-2408. |
| [7] | 王沙沙, 黄超, 汪庆昌, 晁岳恩, 陈锋, 孙建国, 宋晓. 小麦籽粒大小相关基因TaGS2克隆及功能分析[J]. 作物学报, 2022, 48(8): 1926-1937. |
| [8] | 委刚, 陈单阳, 任德勇, 杨宏霞, 伍靖雯, 冯萍, 王楠. 水稻细长秆突变体sr10的鉴定与基因定位[J]. 作物学报, 2022, 48(8): 2125-2133. |
| [9] | 冯亚娟, 李廷轩, 蒲勇, 张锡洲. 不同镉积累类型小麦各器官镉积累分布规律及机理分析[J]. 作物学报, 2022, 48(7): 1761-1770. |
| [10] | 黄福灯, 黄妍, 金泽艳, 贺焕焕, 李春寿, 程方民, 潘刚. 水稻叶片早衰突变体ospls7的生理特性及其基因定位[J]. 作物学报, 2022, 48(7): 1832-1842. |
| [11] | 刘阿康, 马瑞琦, 王德梅, 王艳杰, 杨玉双, 赵广才, 常旭虹. 覆膜和补施氮肥对晚播冬小麦冬前植株生长及群体质量的影响[J]. 作物学报, 2022, 48(7): 1771-1786. |
| [12] | 王娟, 刘翼, 姚丹妤, 邹景伟, 肖世和, 孙果忠. 小麦生殖发育阶段对低温的敏感性鉴定[J]. 作物学报, 2022, 48(7): 1721-1729. |
| [13] | 张少华, 段剑钊, 贺利, 井宇航, 郭天财, 王永华, 冯伟. 基于无人机平台多模态数据融合的小麦产量估算研究[J]. 作物学报, 2022, 48(7): 1746-1760. |
| [14] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [15] | 郑崇珂, 周冠华, 牛淑琳, 和亚男, 孙伟, 谢先芝. 水稻早衰突变体esl-H5的表型鉴定与基因定位[J]. 作物学报, 2022, 48(6): 1389-1400. |
|