欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (11): 2908-2916.doi: 10.3724/SP.J.1006.2024.42013
• 研究简报 • 上一篇
徐飞( ), 刘杨, 许健成, 余璐璐()
), 刘杨, 许健成, 余璐璐()
XU Fei(), LIU Yang, XU Jian-Cheng, YU Lu-Lu()
摘要:
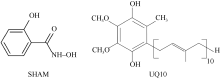
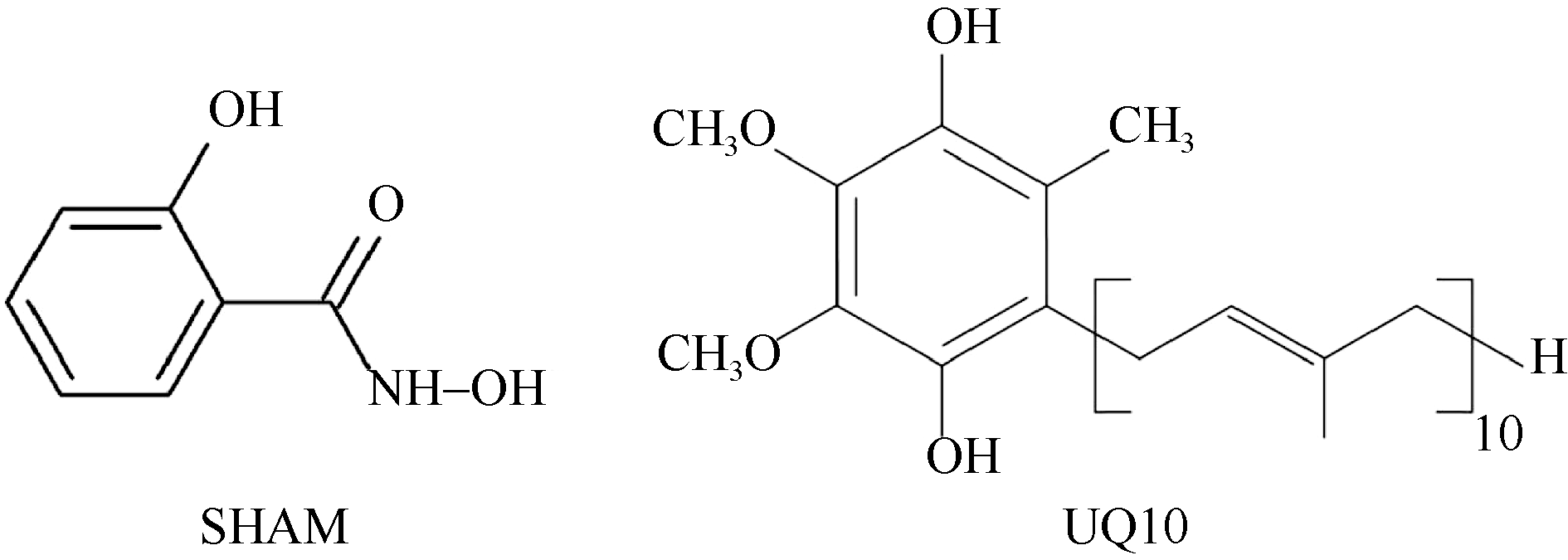
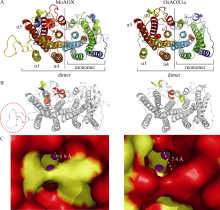
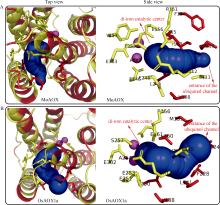
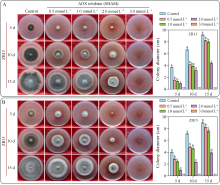
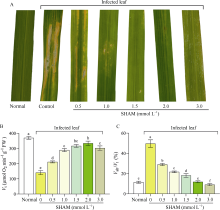
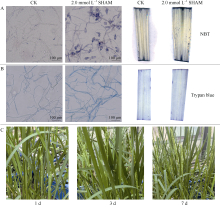
水稻(Oryza sativa L.)是世界主要粮食作物之一。中国作为世界上最大的稻米生产国, 每年因稻瘟菌(Magnaporthe oryzae)引起的稻瘟病受到广泛关注。稻瘟病在水稻整个生育期中均可发生, 从而引起水稻大幅度减产, 甚至颗粒无收。值得注意的是, 交替氧化酶(alternative oxidase, AOX)广泛存在于植物和真菌中, 是线粒体呼吸电子传递链中的末端氧化酶, 被证实参与了胁迫条件下植物的生长发育调节以及真菌胁迫响应等。本文比较分析了水稻AOX和稻瘟菌AOX蛋白结构差异, 并利用AOX抑制剂开展稻瘟菌防治试验。结果表明,水稻AOX和稻瘟菌AOX蛋白的双铁催化活性中心的氨基酸残基相对保守, 但其在泛醌通道大小、氨基酸组成以及形成的疏水口袋等方面有较大差异。通过AOX抑制剂处理可明显减缓稻瘟菌菌丝的生长, 并减轻水稻叶片稻瘟菌病害症状。本研究表明, AOX可作为有效靶标用于抑制稻瘟菌, 在水稻稻瘟菌病害防治方面具有潜在应用价值。
| [1] |
韩艺娟, 鲁国东. 水稻与稻瘟病菌相互作用研究进展. 生物技术通报, 2018, 34(2): 25-37.
doi: 10.13560/j.cnki.biotech.bull.1985.2017-0623 |
| [2] | Han Y J, Lu G D. Recent understanding on the interactions between rice and Magnaporthe oryzae. Biotechnol Bull 2018, 34(2): 25-37 (in Chinese with English abstract). |
| [3] |
Yin J, Zou L, Zhu X, Cao Y, He M, Chen X. Fighting the enemy: How rice survives the blast pathogen’s attack. Crop J, 2021, 9: 543-552.
doi: 10.1016/j.cj.2021.03.009 |
| [4] | Jo L, Pelletier J M, Hsu S W, Baden R, Goldberg R B, Harada J J. Combinatorial interactions of the LEC1 transcription factor specify diverse developmental programs during soybean seed development. Proc Natl Acad Sci USA, 2020, 117: 1223-1232. |
| [5] | Kumar S, Kashyap P L, Mahapatra S, Jasrotia P, Singh G P. New and emerging technologies for detecting Magnaporthe oryzae causing blast disease in crop plants. Crop Prot, 2021, 143: 105473. |
| [6] | Galhano R, Talbot N J. The biology of blast: understanding how Magnaporthe oryzae invades rice plants. Fungal Biol Rev, 2011, 25: 61-67. |
| [7] | Xu F, Copsey A C, Young L, Barsottini M R O, Albury M S, Moore A L. Comparison of the kinetic parameters of alternative oxidases from Trypanosoma brucei and Arabidopsis thaliana: a tale of two cavities. Front Plant Sci, 2021, 12: 744218. |
| [8] |
DelSaz N F, Ribas-Carbo M, McDonald A E, Lambers H, Fernie A R, Florez-Sarasa I. An in vivo perspective of the role(s) of the alternative oxidase pathway. Trends Plant Sci, 2018, 23: 206-219.
doi: S1360-1385(17)30259-5 pmid: 29269217 |
| [9] | Xu F, Peng Y, He Z Q, Yu L L. The role of cyanoalanine synthase and alternative oxidase in promoting salt stress tolerance in Arabidopsis thaliana. BMC Plant Biol 2023, 23: 163. |
| [10] |
Xu F, Yuan S, Zhang D W, Lv X, Lin H H. The role of alternative oxidase in tomato fruit ripening and its regulatory interaction with ethylene. J Exp Bot, 2012, 63: 5705-5716.
doi: 10.1093/jxb/ers226 pmid: 22915749 |
| [11] |
Selinski J, Scheibe R, Day D A, Whelan J. Alternative oxidase is positive for plant performance. Trends Plant Sci, 2018, 23: 588-597.
doi: S1360-1385(18)30080-3 pmid: 29665989 |
| [12] | Moore A L, Albury M S, Crichton P G, Affourtit C. Function of the alternative oxidase: is it still a scavenger. Trends Plant Sci, 2002, 7: 478-481. |
| [13] | Li G B, He J X, Wu J L, Wang H, Zhang X, Liu J, Hu X H, Zhu Y, Shen S, Bai Y F, Yao Z L, Liu X X, Zhao J H, Li D Q, Li Y, Huang F, Huang Y Y, Zhao Z X, Zhang J W, Zhou S X, Ji Y P, Pu M, Qin P, Li S G, Chen X W, Wang J, He M, Li W T, Wu X J, Xu Z J, Wang W M, Fan J. Overproduction of OsRACK1A, an effector-targeted scaffold protein promoting OsRBOHB- mediated ROS production, confers rice floral resistance to false smut disease without yield penalty. Mol Plant, 2022, 15: 1790-1806. |
| [14] |
Li Y, Cao X L, Zhu Y, Yang X M, Zhang K N, Xiao Z Y, Wang H, Zhao J H, Zhang L L, Li G B. Osa-miR398b boosts H2O2 production and rice blast disease-resistance via multiple superoxide dismutases. New Phytol, 2019, 222: 1507-1522.
doi: 10.1111/nph.15678 pmid: 30632163 |
| [15] |
Hu J, Liu M, Zhang A, Dai Y, Chen W, Chen F, Wang W, Shen D, Telebanco-Yanoria M J, Ren B. Co-evolved plant and blast fungus ascorbate oxidases orchestrate the redox state of host apoplast to modulate rice immunity. Mol Plant, 2022, 15: 1347-1366.
doi: 10.1016/j.molp.2022.07.001 pmid: 35799449 |
| [16] | Kido Y, Shiba T, Inaoka D K, Sakamoto K, Nara T, Aoki T, Honma T, Tanaka A, Inoue M, Matsuoka S. Crystallization and preliminary crystallographic analysis of cyanide-insensitive alternative oxidase from Trypanosoma brucei brucei. Acta Crystallogr Sect F Struct Biol Cryst Commun 2010, 66: 275-278. |
| [17] | May B, Young L, Moore A L. Structural insights into the alternative oxidases: are all oxidases made equal. Biochem Soc Trans, 2017, 45: 731-740. |
| [18] |
Kawano Y. Fine-tuning ROS homeostasis by ROD1 is a battleground between rice and Magnaporthe oryzae. Mol Plant 2021, 14: 1979-1981.
doi: 10.1016/j.molp.2021.11.005 pmid: 34785317 |
| [19] | Liu X, Zhang Z. A double-edged sword: reactive oxygen species (ROS) during the rice blast fungus and host interaction. FEBS J, 2022, 289: 5505-5515. |
| [20] | Liu Y, Yu L L, Peng Y, Geng X X, Xu F. Alternative oxidase inhibition impairs tobacco root development and root hair formation. Front Plant Sci, 2021, 12: 664792. |
| [21] | Manbir S P, Kumari A, Gupta K J. Alternative oxidase plays a role in minimizing ROS and RNS produced under salinity stress in Arabidopsis thaliana. Physiol Plant 2022, 174: e13649. |
| [1] | 贾舒涵, 何璨, 陈敏, 刘家欣, 胡伟民, 胡晋, 关亚静. 杂交水稻不同穗萌程度种子质量差异与穗萌分级研究[J]. 作物学报, 2024, 50(9): 2310-2322. |
| [2] | 胡丽琴, 肖正午, 方升亮, 曹放波, 陈佳娜, 黄敏. 种植季节对高直链淀粉水稻品种淀粉消化特性的影响[J]. 作物学报, 2024, 50(9): 2347-2357. |
| [3] | 刘陈, 王昆昆, 廖世鹏, 杨佳群, 丛日环, 任涛, 李小坤, 鲁剑巍. 氮肥用量对玉米-油菜和水稻-油菜轮作模式下油菜产量及氮素吸收利用的影响[J]. 作物学报, 2024, 50(8): 2067-2077. |
| [4] | 宋志文, 赵蕾, 毕俊国, 唐清芸, 王国栋, 李玉祥. 滴灌条件下施氮量对不同氮效率水稻品种物质积累及养分吸收的影响[J]. 作物学报, 2024, 50(8): 2025-2038. |
| [5] | 邵美红, 赵玲玲, 程楚, 程思明, 朱双兵, 翟来圆, 陈凯, 徐建龙. 水稻黄华占背景选择导入系的耐低氮筛选评价与利用[J]. 作物学报, 2024, 50(8): 1907-1919. |
| [6] | 何丹丹, 舒亚洲, 周海连, 吴松果, 魏晓双, 杨明冲, 李波, 吴正丹, 韩世健, 杨娟, 王继斌, 王令强. OsRPTA18参与调控水稻叶片倾角的功能[J]. 作物学报, 2024, 50(8): 1934-1947. |
| [7] | 付景, 马梦娟, 张骐飞, 段居琦, 王越涛, 王付华, 王生轩, 白涛, 尹海庆, 王亚. 干湿交替灌溉和施氮量对粳稻光合特性和氮素吸收利用的影响[J]. 作物学报, 2024, 50(7): 1787-1804. |
| [8] | 裴法敬, 张文轩, 张晓, 王昕钰, 彭少兵, 米甲明. 长粒香型的超短生育期水稻新品系创制[J]. 作物学报, 2024, 50(7): 1684-1698. |
| [9] | 唐清芸, 杨晶晶, 赵蕾, 宋志文, 王国栋, 李玉祥. 施氮量对滴灌水稻根系形态构型和分形特征的影响[J]. 作物学报, 2024, 50(6): 1540-1553. |
| [10] | 张小芳, 朱琪, 华芸堰, 贾黎惠莹, 邱士优, 陈宇杰, 马涛, 丁沃娜. 水稻OsCYP22互作蛋白的筛选及验证[J]. 作物学报, 2024, 50(6): 1628-1634. |
| [11] | 朱忠林, 文月, 周棋, 巫燕飞, 杜雪竹, 盛锋. 水稻OsCNGC10基因抗倒伏性以及抗旱性功能研究[J]. 作物学报, 2024, 50(5): 1351-1360. |
| [12] | 胡明明, 丁峰, 彭志芸, 向开宏, 李郁, 张宇杰, 杨志远, 孙永健, 马均. 多元化种植模式下秸秆还田配合水氮管理对水稻产量形成与氮素吸收利用的影响[J]. 作物学报, 2024, 50(5): 1236-1252. |
| [13] | 耿孝宇, 张翔, 刘洋, 左博源, 朱旺, 马唯一, 汪璐璐, 孟天瑶, 高平磊, 陈英龙, 许轲, 戴其根, 韦还和. 江苏省滨海盐碱地籼粳杂交稻产量优势形成特征[J]. 作物学报, 2024, 50(5): 1253-1270. |
| [14] | 万应春, 班义结, 蒋钰东, 王亚欣, 刘晶晶, 刘晓晴, 程育林, 王楠, 冯萍. 水稻雄性不育突变体tpa1的表型鉴定与精细定位[J]. 作物学报, 2024, 50(5): 1104-1114. |
| [15] | 余瑶, 王紫瑶, 周思睿, 刘鹏程, 叶亚峰, 马伯军, 刘斌美, 陈析丰. 水稻类病变突变体lms1的表型鉴定与抗病分子机制分析[J]. 作物学报, 2024, 50(4): 857-870. |
|
||