欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (5): 1124-1135.doi: 10.3724/SP.J.1006.2024.33044
苏帅( ), 刘孝伟, 牛群凯, 时子文, 侯雨微, 冯开洁, 荣廷昭, 曹墨菊*()
), 刘孝伟, 牛群凯, 时子文, 侯雨微, 冯开洁, 荣廷昭, 曹墨菊*()
SU Shuai(), LIU Xiao-Wei, NIU Qun-Kai, SHI Zi-Wen, HOU Yu-Wei, FENG Kai-Jie, RONG Ting-Zhao, CAO Mo-Ju*()
摘要:
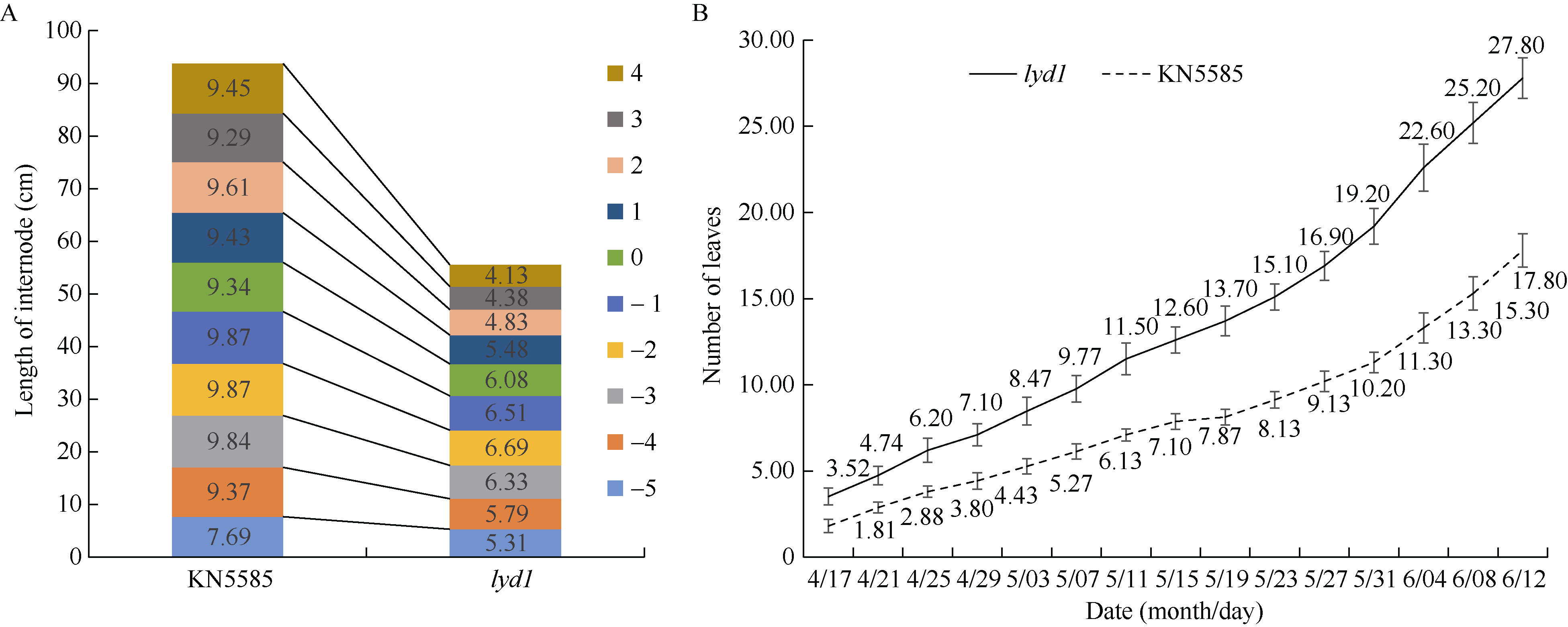
玉米株高降低通常是由节间数目减少、节间长度变短或二者共同作用所致。而本研究在基因编辑后代中发现的玉米多叶矮化突变体lyd1却表现为节间数目显著增加, 株高显著降低。lyd1株高仅为93.10 cm, 与野生型KN5585的株高159.95 cm相比, 降低了41.79%, 差异达到极显著水平。然而lyd1叶片数平均达到27.8片, 相较野生型平均17.8片叶, 增加56.18%, 差异达到极显著水平。遗传分析表明, lyd1的突变表型由1对隐性核基因控制, 通过图位克隆将控制多叶矮化性状的基因定位于玉米3号染色体标记Indel10和Indel11之间, 物理距离0.74 Mb。对定位区间内13个基因(不包含假基因)的序列进行测序, 发现仅ZmTE1在第4外显子出现1个A碱基的替换, 其他基因无差异。ZmTE1编码一个RNA结合蛋白, 氨基酸的替换发生在第3个RNA结合结构域内(RRM3), 导致天冬氨酸转变为缬氨酸。突变体lyd1的突变位点与已报道的te1-mum1、te1-mum2、te1-mum3、zm66不同, lyd1的发现为进一步解析玉米叶片和节间发育平衡的遗传机制提供了宝贵的材料。
| [1] |
Tester M, Langridge P. Breeding technologies to increase crop production in a changing world. Science, 2010, 327: 818-822.
doi: 10.1126/science.1183700 pmid: 20150489 |
| [2] | Haarhoff S J, Swanepoel P A. Plant population and maize grain yield: a global systematic review of rainfed trials. Crop Sci, 2018, 5: 1819-1829. |
| [3] |
Bensen R J, Johal G S. Cloning and characterization of the maize AN1 gene. Plant Cell, 1995, 7: 75-84.
doi: 10.1105/tpc.7.1.75 pmid: 7696880 |
| [4] |
Chen Y, Hou M M, Liu L J, Wu S, Shen Y, Ishiyama K, Kobaya, Shi M, McCarty D R, Tan B C. The maize Dwarf1 encodes a gibberellin 3-oxidase and is dual localized to the nucleus and cytosol. Plant Physiol, 2014, 166: 2028-2039.
doi: 10.1104/pp.114.247486 pmid: 25341533 |
| [5] |
Teng F, Zhai L H, Liu R X, Bai W, Wang L Q, Huo D G, Tao Y S, Zheng Y L, Zhang Z X. ZmGA3ox2, a candidate gene for a major QTL, qPH3.1, for plant height in maize. Plant J, 2013, 73: 405-416.
doi: 10.1111/tpj.2013.73.issue-3 |
| [6] |
Winkler R G, Helentjaris T. The maize Dwarf3 gene encodes a cytochrome P450-mediated early step in gibberellin biosynthesis. Plant Cell, 1995, 7: 1307-1317.
doi: 10.1105/tpc.7.8.1307 pmid: 7549486 |
| [7] |
Cassani E, Bertolini E, Cerino Badone F, Landoni M, Gavina D, Sirizzotti A, Pilu R. Characterization of the first dominant dwarf maize mutant carrying a single amino acid insertion in the VHYNP domain of the Dwarf8 gene. Mol Breed, 2009, 24: 375-385.
doi: 10.1007/s11032-009-9298-3 |
| [8] |
Harberd N P, Freeling M. Genetics of dominant gibberellin- insensitive dwarfism in maize. Genetics, 1989, 121: 827-838.
doi: 10.1093/genetics/121.4.827 pmid: 17246493 |
| [9] |
Lawit S J, Wych H M, Xu D, Kundu S, Tomes D T. Maize DELLA proteins Dwarf plant8 and Dwarf plant9 as modulators of plant development. Plant Cell Physiol, 2010, 51: 1854-1868.
doi: 10.1093/pcp/pcq153 |
| [10] |
Multani D S, Briggs S P, Chamberlin M A, Blakeslee J J, Murphy A S, Johal G S. Loss of an MDR transporter in compact stalks of maize Br2and sorghum Dw3 mutants. Science, 2003, 302: 81-84.
doi: 10.1126/science.1086072 pmid: 14526073 |
| [11] |
Zhang X, Hou X, Liu Y, Zheng L, Yi Q, Zhang H, Huang X, Zhang J, Hu Y, Yu G, Liu H, Li Y, Huang H, Zhan F, Chen L, Tang J, Huang Y. Maize brachytic2 (Br2) suppresses the elongation of lower internodes for excessive auxin accumulation in the intercalary meristem region. BMC Plant Biol, 2019, 19: 589.
doi: 10.1186/s12870-019-2200-5 pmid: 31881837 |
| [12] |
Hartwig T, Chuck G S, Fujioka S, Klempien A, Weizbauer R, Potluri D P V, Choe S, Johal G S, Schulz B. Brassinosteroid control of sex determination in maize. Proc Natl Acad Sci USA, 2011, 108: 19814-19819.
doi: 10.1073/pnas.1108359108 pmid: 22106275 |
| [13] |
Best N B, Hartwig T, Budka J, Fujioka S, Johal G, Schulz B, Dilkes B P. Nana plant2 encodes a maize ortholog of the Arabidopsis brassinosteroid biosynthesis gened Dwarf1, identifying developmental interactions between brassinosteroids and gibberellins. Plant Physiol, 2016, 171: 2633-2647.
doi: 10.1104/pp.16.00399 |
| [14] | Makarevitch I, Thompson A, Muehlbauer G J, Springer N M. Brd1 gene in maize encodes a brassinosteroid C-6 oxidase. PLoS One, 2012, 7: e30798. |
| [15] |
Kir G, Ye H, Nelissen H, Neelakandan A K, Kusnandar A S, Luo A, Inzé D, Sylvester A W, Yin Y, Becraft P W. RNA interference knock down of BRI1 in maize reveals novel functions for brassinosteroid signaling in controlling plant architecture. Plant Physiol, 2015, 169: 826-839.
doi: 10.1104/pp.15.00367 |
| [16] |
Li H, Wang L, Liu M, Dong Z, Li Q, Fei S, Xiang H, Liu B, Jin W. Maize plant architecture is regulated by the ethylene biosynthetic gene ZmACS7. Plant Physiol, 2020, 183: 1184-1199.
doi: 10.1104/pp.19.01421 |
| [17] |
Schaller G E, Bishopp A, Kieber J J. The Yin-Yang of hormones: cytokinin and auxin interactions in plant development. Plant Cell, 2015, 27: 44-63.
doi: 10.1105/tpc.114.133595 |
| [18] |
Phillips K A, Skirpan A L, Liu X, Christensen A, Slewinski T L, Hudson C, Barazesh S, Cohen J D, Malcomber S, Mcsteen P. Vanishing tassel2 encodes a grass-specific tryptophan aminotransferase required for vegetative and reproductive development in maize. Plant Cell, 2011, 23: 550-566.
doi: 10.1105/tpc.110.075267 |
| [19] | Lee B H, Johnston R, Yang Y, Gallavotti A, Kojima M, Travençolo B A, Costa Lda F, Sakakibara H, Jackson D. Studies of Aberrant phyllotaxy1 mutants of maize indicate complex interactions between auxin and cytokinin signaling in the shoot apical meristem. Plant Physiol, 2009, 150: 205-216. |
| [20] |
张在宝, 李婉杰, 李九丽, 张弛, 胡梦辉, 程琳, 袁红雨. 植物RNA结合蛋白研究进展. 中国农业科学, 2018, 51: 4007-4019.
doi: 10.3864/j.issn.0578-1752.2018.21.001 |
|
Zhang Z B, Li W J, Li J L, Zhang C, Hu M H, Cheng L, Yuan H Y. The research progress of plant RNA binding proteins. Sci Agric Sin, 2018, 51: 4007-4019 (in Chinese with English abstract).
doi: 10.3864/j.issn.0578-1752.2018.21.001 |
|
| [21] |
Cho H, Cho H S, Hwang I. Emerging roles of RNA-binding proteins in plant development. Curr Opin Plant Biol, 2019, 51: 51-57.
doi: S1369-5266(19)30026-3 pmid: 31071564 |
| [22] |
Jeffares D C, Phillips M J, Moore S, Veit B. A description of the Mei2-like protein family; structure, phylogenetic distribution and biological context. Dev Genes Evol, 2004, 214: 149-158.
pmid: 14986133 |
| [23] |
Kawakatsu T, Itoh J, Miyoshi K, Kurata N, Alvarez N, Veit B, Nagato Y. PLASTOCHRON2 regulates leaf initiation and maturation in rice. Plant Cell, 2006, 18: 612-625.
pmid: 16461585 |
| [24] |
Anderson G H, Alvarez N D, Gilman C, Jeffares D C, Trainor V C, Hanson M R, Veit B. Diversification of genes encoding mei2-like RNA binding proteins in plants. Plant Mol Biol, 2004, 54: 653-670.
pmid: 15356386 |
| [25] | 王关林, 方宏筠. 植物基因工程(第2版). 北京: 科学出版社, 2002. pp 742-744. |
| Wang G L, Fang H Y. Plant Gene Engineering, 2nd edn. Beijing: Science Press, 2002. pp 742-744 (in Chinese). | |
| [26] |
Michelmore R W, Paran I, Kesseli R V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832.
doi: 10.1073/pnas.88.21.9828 pmid: 1682921 |
| [27] |
Avila L M, Cerrudo D, Swanton C, Lukens L. Brevis plant1, a putative inositol polyphosphate 5-phosphatase, is required for internode elongation in maize. J Exp Bot, 2016, 67: 1577-1588.
doi: 10.1093/jxb/erv554 pmid: 26767748 |
| [28] |
Zhang D, Sun W, Singh R, Zheng Y, Cao Z, Li M, Lunde C, Hake S, Zhang Z. GRF-interacting factor1 regulates shoot architecture and meristem determinacy in maize. Plant Cell, 2018, 30: 360-374.
doi: 10.1105/tpc.17.00791 |
| [29] |
Li W, Ge F, Qiang Z, Zhu L, Zhang S, Chen L, Wang X, Li J, Fu Y. Maize ZmRPH1 encodes a microtubule-associated protein that controls plant and ear height. Plant Biotechnol J, 2020, 18: 1345-1347.
doi: 10.1111/pbi.v18.6 |
| [30] |
Heuer S, Hansen S, Bantin J, Brettschneider R, Kranz E, Lörz H, Dresselhaus T. The maize MADS box gene ZmMADS3 affects node number and spikelet development and is co-expressed with ZmMADS1 during flower development, in egg cells, and early embryogenesis. Plant Physiol, 2001, 127: 33-45.
doi: 10.1104/pp.127.1.33 pmid: 11553732 |
| [31] |
Lyu H K, Zheng J, Wang T Y, Fu J J, Huai J L, Min H W, Zhang X, Tian B H, Shi Y S, Wang G Y. The maize d2003, a novel allele of VP8, is required for maize internode elongation. Plant Mol Biol, 2014, 84: 243-257.
doi: 10.1007/s11103-013-0129-x |
| [32] |
Bommert P, Je B I, Goldshmidt A, Jackson D. The maize Gα gene Compact plant2 functions in clavata signalling to control shoot meristem size. Nature, 2013, 502: 555-558.
doi: 10.1038/nature12583 |
| [33] |
Veit B, Briggs S P. Regulation of leaf initiation by the terminal ear1 gene of maize. Nature, 1998, 393: 166-168.
doi: 10.1038/30239 |
| [34] |
Wang F, Yu Z, Zhang M, Wang M, Lu X, Liu X, Li Y, Zhang X, Tan B C, Li C, Ding Z. ZmTE1 promotes plant height by regulating intercalary meristem formation and internode cell elongation in maize. Plant Biotechnol J, 2022, 20: 526-537.
doi: 10.1111/pbi.v20.3 |
| [35] |
Hentze M W, Castello A, Schwarzl T, Preiss T. A brave new world of RNA-binding proteins. Nat Rev Mol Cell Biol, 2018, 19: 327-341.
doi: 10.1038/nrm.2017.130 |
| [36] | 唐蜻. 植物RNA结合蛋白的研究进展. 安徽农业科学, 2010, 38(1): 38-41. |
| Tang Q. The research progress of plant RNA binding proteins. J Anhui Agric Sci, 2010, 38(1): 38-41 (in Chinese with English abstract). |
| [1] | 王菲儿, 郭瑶, 李盼, 韦金贵, 樊志龙, 胡发龙, 范虹, 何蔚, 殷文, 陈桂平. 绿洲灌区增密对水氮减量玉米产量的补偿机制[J]. 作物学报, 2024, 50(6): 1616-1627. |
| [2] | 折萌, 郑登俞, 柯照, 吴忠义, 邹华文, 张中保. 玉米ZmGRAS13基因的克隆及功能研究[J]. 作物学报, 2024, 50(6): 1420-1434. |
| [3] | 郑雪晴, 王兴荣, 张彦军, 龚佃明, 邱法展. 玉米果穗相关性状QTL定位及重要候选基因分析[J]. 作物学报, 2024, 50(6): 1435-1450. |
| [4] | 韩洁楠, 张泽, 刘晓丽, 李冉, 上官小川, 周婷芳, 潘越, 郝转芳, 翁建峰, 雍洪军, 周志强, 徐晶宇, 李新海, 李明顺. o2突变引起糯玉米籽粒淀粉积累差异研究[J]. 作物学报, 2024, 50(5): 1207-1222. |
| [5] | 王永亮, 胥子航, 李申, 梁哲铭, 白炬, 杨治平. 不同覆盖措施对土壤水热状况及春玉米产量和水分利用效率的影响[J]. 作物学报, 2024, 50(5): 1312-1324. |
| [6] | 田红丽, 杨扬, 范亚明, 易红梅, 王蕊, 金石桥, 晋芳, 张云龙, 刘亚维, 王凤格, 赵久然. 用于玉米品种真实性鉴定的最优核心SNP位点集的研发[J]. 作物学报, 2024, 50(5): 1115-1123. |
| [7] | 余瑶, 王紫瑶, 周思睿, 刘鹏程, 叶亚峰, 马伯军, 刘斌美, 陈析丰. 水稻类病变突变体lms1的表型鉴定与抗病分子机制分析[J]. 作物学报, 2024, 50(4): 857-870. |
| [8] | 邹佳琪, 王仲林, 谭先明, 陈燎原, 杨文钰, 杨峰. 基于连续小波变换估测干旱胁迫下玉米籽粒产量[J]. 作物学报, 2024, 50(4): 1030-1042. |
| [9] | 娄菲, 左怿平, 李萌, 代鑫萌, 王健, 韩金玲, 吴舒, 李向岭, 段会军. 有机肥替代部分化肥氮对糯玉米产量、品质及氮素利用的影响[J]. 作物学报, 2024, 50(4): 1053-1064. |
| [10] | 岳海旺, 魏建伟, 刘朋程, 陈淑萍, 卜俊周. 基于GYT双标图分析对黄淮海生态区玉米品种综合评价[J]. 作物学报, 2024, 50(4): 836-856. |
| [11] | 薛明, 汪晨晨, 姜露光, 刘浩, 张路遥, 陈赛华. 玉米花序发育基因AFP1的定位及功能研究[J]. 作物学报, 2024, 50(3): 603-612. |
| [12] | 赵荣荣, 丛楠, 赵闯. 基于Landsat 8影像提取豫中地区冬小麦和夏玉米分布信息的最佳时相选择[J]. 作物学报, 2024, 50(3): 721-733. |
| [13] | 梁星伟, 杨文亭, 金雨, 胡莉, 傅小香, 陈先敏, 周顺利, 申思, 梁效贵. 玉米穗轴的颜色变化, 是偶然还是与农艺性状存在关联?——以历年国审普通品种为例[J]. 作物学报, 2024, 50(3): 771-778. |
| [14] | 毛燕, 郑名敏, 牟成香, 谢吴兵, 唐琦. 渗透胁迫下玉米自然反义转录本cis-NATZmNAC48启动子的功能分析[J]. 作物学报, 2024, 50(2): 354-362. |
| [15] | 马娟, 曹言勇. 玉米杂交群体产量性状及其特殊配合力全基因组关联分析[J]. 作物学报, 2024, 50(2): 363-372. |
|