欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (12): 3013-3024.doi: 10.3724/SP.J.1006.2024.44060
方然1( ), 袁丽媚1, 王玉林1, 芦思佳1,2, 孔凡江1,2, 刘宝辉1,2,*(), 孔令平1,2,*()
), 袁丽媚1, 王玉林1, 芦思佳1,2, 孔凡江1,2, 刘宝辉1,2,*(), 孔令平1,2,*()
FANG Ran1(), YUAN Li-Mei1, WANG Yu-Lin1, LU Si-Jia1,2, KONG Fan-Jiang1,2, LIU Bao-Hui1,2,*(), KONG Ling-Ping1,2,*()
摘要:
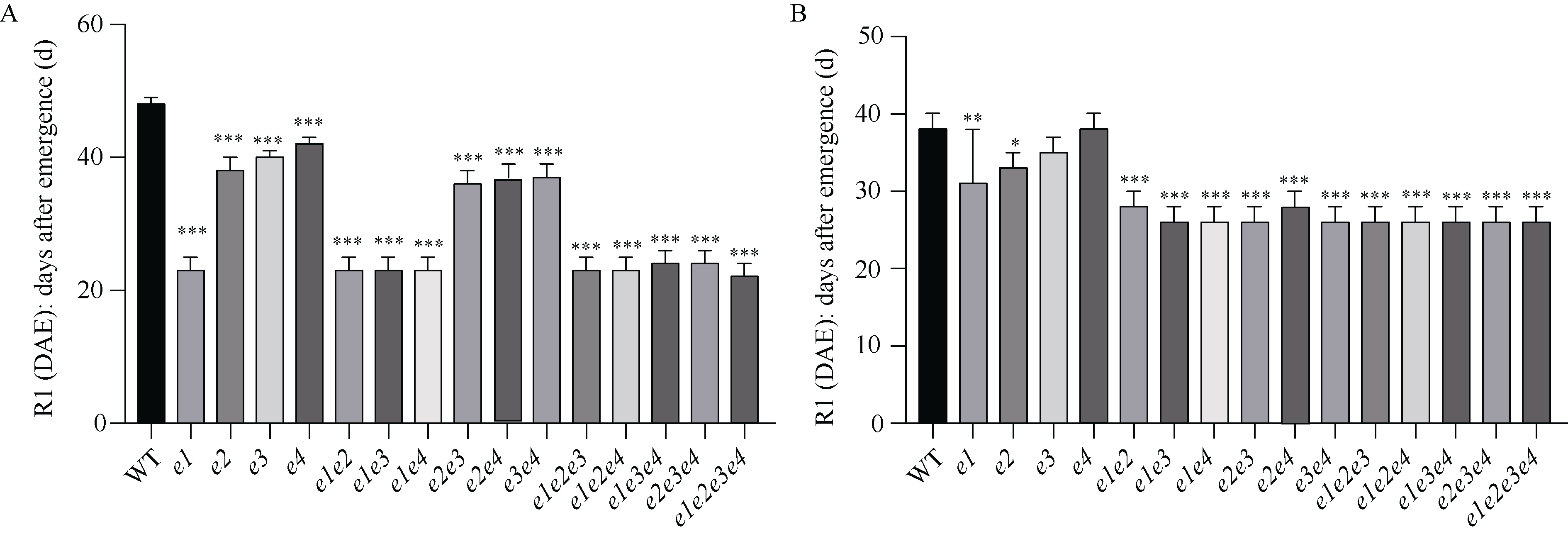
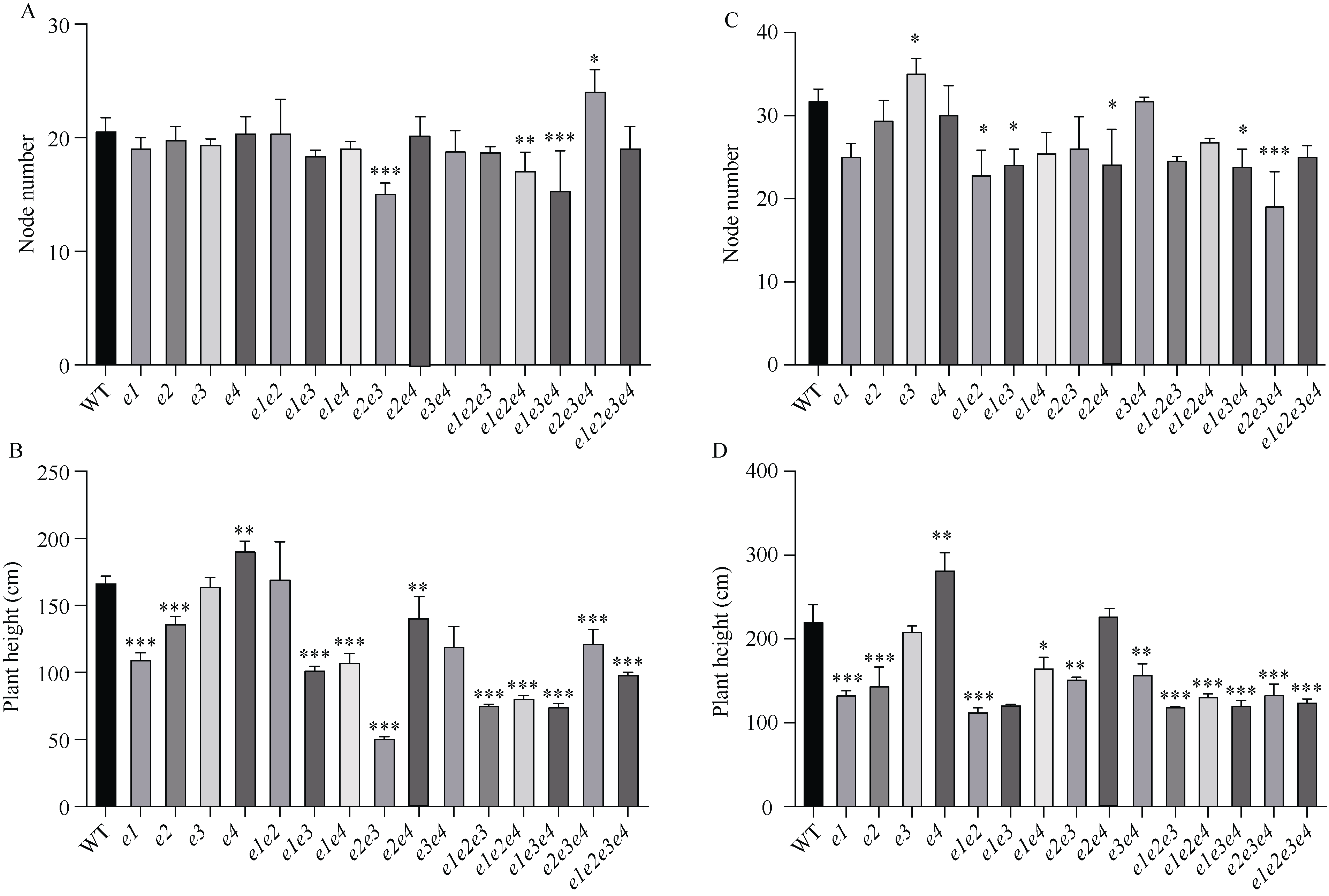
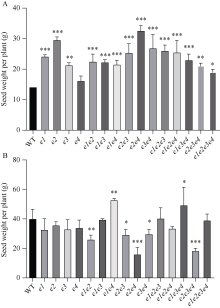
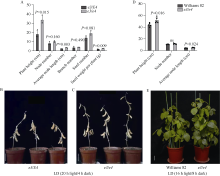
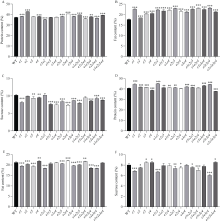
大豆作为重要的油料作物, 是人类优质蛋白和食用油的主要来源之一。大豆产量、种子品质和生育期性状密切相关, 生育期性状主要受一系列生育期相关基因的控制。本研究以Harosoy为遗传背景创制了E1~E4不同基因组合的16种近等基因系材料, 种植在河北石家庄和安徽合肥试验田, 调查了生育期、品质及产量性状, 以此来了解E1~E4不同突变组合对中纬度种植区域的适应性。研究结果表明, 16种近等基因系材料, 对光周期敏感程度不同, 开花期存在差异, WT和e4单突近等基因系由于晚花晚熟、产量低, 不适合在石家庄种植, 而所有的近等基因系在合肥种植时均能正常成熟。E1~E4不同等位基因组合还影响株高、节间距、单株产量、品质等农艺性状。本研究发现, 在长日照条件下, e3或e4单独突变可以提前开花, 同时还会使大豆产生避荫反应, 株高增高、节间距变大。测量种子的蛋白质、油分和蔗糖含量时发现, WT材料在较高纬度的石家庄地区种植时, 种子不能正常成熟, 含油量最低, 而蔗糖的含量最高。整体来看, 其余近等基因系的种子在石家庄地区种植时总油分、蔗糖含量高于合肥, 而蛋白含量总体低于合肥地区。因此, 在评估大豆品种的纬度适应性时, 需综合考察生育期基因对光周期敏感性、品质和产量的影响。
| [1] |
Cao D, Takeshima R, Zhao C, Liu B H, Jun A, Kong F J. Molecular mechanisms of flowering under long days and stem growth habit in soybean. J Exp Bot, 2017, 68: 1873-1884.
doi: 10.1093/jxb/erw394 pmid: 28338712 |
| [2] |
Lin X Y, Fang C, Liu B H, Kong F J. Natural variation and artificial selection of photoperiodic flowering genes and their applications in crop adaptation. aBIOTECH, 2021, 2: 156-169.
doi: 10.1007/s42994-021-00039-0 pmid: 36304754 |
| [3] | Destro D, Carpentieri-Pípolo V, Kiihl R A S, Almeida L A. Photoperiodism and genetic control of the long juvenile period in soybean: a review. Crop Breed Appl Biotechnol, 2001, 1: 72-92. |
| [4] | Mourtzinis S, Conley S P. Delineating soybean maturity groups across the United States. Agron J, 2017, 109: 1397-1403. |
| [5] | Jia H C, Jiang B J, Wu C X, Lu W C, Hou W S, Sun S, Yan H R, Han T F. Maturity group classification and maturity locus genotyping of early-maturing soybean varieties from high-latitude cold regions. PLoS One, 2014, 9: e94139. |
| [6] | Li J C, Wang X B, Song W W, Huang X Y, Zhou J, Zeng H Y, Sun S, Jia H C, Li W B, Zhou X N, Li S Z, Chen P Y, Wu C X, Guo Y, Han T F, Qiu L J. Genetic variation of maturity groups and four E genes in the Chinese soybean mini core collection. PLoS One, 2017, 12: e0172106. |
| [7] | Yang W Y, Wu T T, Zhang X Y, Song W W, Xu C L, Sun S, Hou W S, Jiang B J, Han T F, Wu C X. Critical photoperiod measurement of soybean genotypes in different maturity groups. Crop Sci, 2019, 59: 2055-2061. |
| [8] | Xia Z J, Watanabe S, Yamada T, Tsubokura Y, Nakashima H, Zhai H, Anai T, Sato S, Yamazaki T, Lyu S X, Wu H Y, Tabata S, Harada K. Positional cloning and characterization reveal the molecular basis for soybean maturity locus E1 that regulates photoperiodic flowering. Proc Natl Acad Sci USA, 2012, 109: E2155-E2164. |
| [9] |
Xu M L, Yamagishi N, Zhao C, Takeshima R, Kasai M, Watanabe S, Kanazawa A, Yoshikawa N, Liu B H, Yamada T, Abe J. The soybean-specific maturity gene E1 family of floral repressors controls night-break responses through down-regulation of FLOWERING LOCUS T orthologs. Plant Physiol, 2015, 168: 1735-1746.
doi: 10.1104/pp.15.00763 |
| [10] |
Watanabe S, Xia Z J, Hideshima R, Tsubokura Y, Sato S, Yamanaka N, Takahashi R, Anai T, Tabata S, Kitamura K, Harada K. A map-based cloning strategy employing a residual heterozygous line reveals that the GIGANTEA gene is involved in soybean maturity and flowering. Genetics, 2011, 188: 395-407.
doi: 10.1534/genetics.110.125062 pmid: 21406680 |
| [11] |
Watanabe S, Hideshima R, Xia Z J, Tsubokura Y, Sato S, Nakamoto Y, Yamanaka N, Takahashi R, Ishimoto M, Anai T, Tabata S, Harada K. Map-based cloning of the gene associated with the soybean maturity locus E3. Genetics, 2009, 182: 1251-1262.
doi: 10.1534/genetics.108.098772 pmid: 19474204 |
| [12] |
Liu B H, Kanazawa A, Matsumura H, Takahashi R, Harada K, Abe J. Genetic redundancy in soybean photoresponses associated with duplication of the phytochrome A gene. Genetics, 2008, 180: 995-1007.
doi: 10.1534/genetics.108.092742 pmid: 18780733 |
| [13] | Cober E R. Long juvenile soybean flowering responses under very short photoperiods. Crop Sci, 2011, 51: 140-145. |
| [14] | Cober E R, Voldeng H D. Low R: FR light quality delays flowering of E7E7 soybean lines. Crop Sci, 2001, 41: 1823-1826. |
| [15] | Cober E R, Molnar S J, Charette M, Voldeng H D. A new locus for early maturity in soybean. Crop Sci, 2010, 50: 524-527. |
| [16] | Kong F J, Nan H Y, Cao D, Li Y, Wu F F, Wang J L, Lu S J, Yuan X H, Cober E R, Abe J, Liu B H. A new dominant gene E9 conditions early flowering and maturity in soybean. Crop Sci, 2014, 54: 2529-2535. |
| [17] | Kong F J, Liu B H, Xia Z J, Sato S, Kim B M, Watanabe S, Yamada T, Tabata S, Kanazawa A, Harada K, Abe J. Two coordinately regulated homologs of FLOWERING LOCUS T are involved in the control of photoperiodic flowering in soybean. Plant Physiol, 2010, 154: 1220-1231. |
| [18] |
Samanfar B, Molnar S J, Charette M, Schoenrock A, Dehne F, Golshani A, Belzile F, Cober E R. Mapping and identification of a potential candidate gene for a novel maturity locus, E10, in soybean. Theor Appl Genet, 2017, 130: 377-390.
doi: 10.1007/s00122-016-2819-7 pmid: 27832313 |
| [19] | Lu S J, Dong L D, Fang C, Liu S L, Kong L P, Cheng Q, Chen L Y, Su T, Nan H Y, Zhang D, Zhang L, Wang Z J, Yang Y Q, Yu D Y, Liu X L, Yang Q Y, Lin X Y, Tang Y, Zhao X H, Yang X Q, Tian C G, Xie Q G, Li X, Yuan X H, Tian Z X, Liu B H, Weller J L, Kong F J. Stepwise selection on homeologous PRR genes controlling flowering and maturity during soybean domestication. Nat Genet, 2020, 52: 428-436. |
| [20] | 侯智红. 大豆高纬度适应性相关位点Tof5的克隆与功能研究. 黑龙江八一农垦大学博士学位论文,黑龙江大庆, 2022. |
| Hou Z H. Cloning and Functional Analysis of Tof5, a Locus of Soybean Adaptated to High Latitude. PhD Dissertation of Heilongjiang Bayi Agricultural University, Daqing, Heilongjiang, China, 2022 (in Chinese with English abstract). | |
| [21] |
Lu S J, Zhao X H, Hu Y L, Liu S L, Nan H Y, Li X M, Fang C, Cao D, Shi X Y, Kong L P, Su T, Zhang F G, Li S C, Wang Z, Yuan X H, Cober E R, Weller J L, Liu B H, Hou X L, Tian Z X, Kong F J. Natural variation at the soybean J locus improves adaptation to the tropics and enhances yield. Nat Genet, 2017, 49: 773-779.
doi: 10.1038/ng.3819 |
| [22] | Lu S J, Li Y, Wang J L, Srinives P, Nan H Y, Cao D, Wang Y P, Li J L, Li X M, Fang C, Shi X Y, Yuan X H, Watanabe S, Feng X Z, Liu B H, Abe J, Kong F J. QTL mapping for flowering time in different latitude in soybean. Euphytica, 2015, 206: 725-736. |
| [23] | Lin X Y, Dong L D, Tang Y, Li H Y, Cheng Q, Li H, Zhang T, Ma L X, Xiang H L, Chen L N, Nan H Y, Fang C, Lu S J, Li J G, Liu B H, Kong F J. Novel and multifaceted regulations of photoperiodic flowering by phytochrome A in soybean. Proc Natl Acad Sci USA, 2022, 119: e2208708119. |
| [24] | Li H Y, Du H P, He M L, Wang J H, Wang F, Yuan W J, Huang Z R, Cheng Q, Gou C J, Chen Z, Liu B H, Kong F J, Fang C, Zhao X H, Yu D Y. Natural variation of FKF1 controls flowering and adaptation during soybean domestication and improvement. New Phytol, 2023, 238: 1671-1684. |
| [25] | Dong L D, Cheng Q, Fang C, Kong L P, Yang H, Hou Z H, Li Y L, Nan H Y, Zhang Y H, Chen Q S, Zhang C B, Kou K, Su T, Wang L S, Li S C, Li H Y, Lin X Y, Tang Y, Zhao X H, Lu S J, Liu B H, Kong F J. Parallel selection of distinct Tof5 alleles drove the adaptation of cultivated and wild soybean to high latitudes. Mol Plant, 2022, 15: 308-321. |
| [26] | Bonato E R, Vello N A. E6, a dominant gene conditioning early flowering and maturity in soybeans. Genet Mol Biol, 1999, 22: 229-232. |
| [27] | Ray J D, Hinson K, Mankono J E B, Malo M F. Genetic control of a long-juvenile trait in soybean. Crop Sci, 1995, 35: 1001-1006. |
| [28] | Fang C, Liu J, Zhang T, Su T, Li S C, Cheng Q, Kong L P, Li X M, Bu T T, Li H Y, Dong L D, Lu S J, Kong F J, Liu B H. A recent retrotransposon insertion of J caused E6 locus facilitating soybean adaptation into low latitude. J Integr Plant Biol, 2021, 63: 995-1003. |
| [29] |
Dong L D, Fang C, Cheng Q, Su T, Kou K, Kong L P, Zhang C B, Li H Y, Hou Z H, Zhang Y H, Chen L Y, Yue L, Wang L S, Wang K, Li Y L, Gan Z R, Yuan X H, Weller J L, Lu S J, Kong F J, Liu B H. Genetic basis and adaptation trajectory of soybean from its temperate origin to tropics. Nat Commun, 2021, 12: 5445.
doi: 10.1038/s41467-021-25800-3 pmid: 34521854 |
| [30] | Fehr W R, Caviness C E, Burmood D T, Pennington J S. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci, 1971, 11: 929-931. |
| [31] | Tsubokura Y, Watanabe S, Xia Z J, Kanamori H, Yamagata H, Kaga A, Katayose Y, Abe J, Ishimoto M, Harada K. Natural variation in the genes responsible for maturity loci E1, E2, E3 and E4 in soybean. Ann Bot, 2014, 113: 429-441. |
| [32] | Krezhova D. Soybean - Genetics and Novel Techniques for Yield Enhancement. Internet: InTechOpen, 2011. pp 51-76. |
| [33] |
Xu M L, Xu Z H, Liu B H, Kong F J, Tsubokura Y, Watanabe S, Xia Z J, Harada K, Kanazawa A, Yamada T, Abe J. Genetic variation in four maturity genes affects photoperiod insensitivity and PHYA-regulated post-flowering responses of soybean. BMC Plant Biol, 2013, 13: 91.
doi: 10.1186/1471-2229-13-91 pmid: 23799885 |
| [34] | Tsubokura Y, Matsumura H, Xu M L, Liu B H, Nakashima H, Anai T, Kong F J, Yuan X H, Kanamori H, Katayose Y, Takahashi R, Harada K, Abe J. Genetic variation in soybean at the maturity locus E4 Is involved in adaptation to long days at high latitudes. Agronomy, 2013, 3: 117-134. |
| [35] | Abe J, Xu D H, Miyano A, Komatsu K, Kanazawa A, Shimamoto Y. Photoperiod-insensitive Japanese soybean landraces differ at two maturity loci. Crop Sci, 2003, 43: 1300-1304. |
| [36] |
Liu B H, Abe J. QTL mapping for photoperiod insensitivity of a Japanese soybean landrace Sakamotowase. J Hered, 2010, 101: 251-256.
doi: 10.1093/jhered/esp113 pmid: 19959597 |
| [37] | Cober E R, Tanner J W, Voldeng H D. Genetic control of photoperiod response in early-maturing, near-isogenic soybean lines. Crop Sci, 1996, 36: 601-605. |
| [38] |
Cober E R, Morrison M J. Regulation of seed yield and agronomic characters by photoperiod sensitivity and growth habit genes in soybean. Theor Appl Genet, 2010, 120: 1005-1012.
doi: 10.1007/s00122-009-1228-6 pmid: 20012856 |
| [39] |
Fang C, Ma Y M, Wu S W, Liu Z, Wang Z, Yang R, Hu G H, Zhou Z K, Yu H, Zhang M, Pan Y, Zhou G A, Ren H X, Du W G, Yan H R, Wang Y P, Han D Z, Shen Y T, Liu S L, Liu T F, Zhang J X, Qin H, Yuan J, Yuan X H, Kong F J, Liu B H, Li J Y, Zhang Z W, Wang G D, Zhu B G, Tian Z X. Genome-wide association studies dissect the genetic networks underlying agronomical traits in soybean. Genome Biol, 2017, 18: 161.
doi: 10.1186/s13059-017-1289-9 pmid: 28838319 |
| [40] | Wan Z, Liu Y X, Guo D D, Fan R, Liu Y, Xu K, Zhu J L, Quan L, Lu W T, Bai X, Zhai H. CRISPR/Cas9-mediated targeted mutation of the E1 decreases photoperiod sensitivity, alters stem growth habits, and decreases branch number in soybean. Front Plant Sci, 2022, 13: 1066820. |
| [41] | Carrera C, Martínez M J, Dardanelli J, Balzarini M. Environmental variation and correlation of seed components in nontransgenic soybeans: protein, oil, unsaturated fatty acids, tocopherols, and isoflavones. Crop Sci, 2011, 51: 800-809. |
| [42] | Song W W, Yang R P, Wu T T, Wu C X, Sun S, Zhang S W, Jiang B J, Tian S Y, Liu X B, Han T F. Analyzing the effects of climate factors on soybean protein, oil contents, and composition by extensive and high-density sampling in China. J Agric Food Chem, 2016, 64: 4121-4130. |
| [43] |
Song W W, Yang R P, Yang X S, Sun S, Mentreddy S R, Jiang B J, Wu T T, Tian S Y, Sapey E, Wu C X, Hou W S, Ren G X, Han T F. Spatial differences in soybean bioactive components across China and their influence by weather factors. Crop J, 2018, 6: 659-668.
doi: 10.1016/j.cj.2018.05.001 |
| [1] | 赵黎明, 段绍彪, 项洪涛, 郑殿峰, 冯乃杰, 沈雪峰. 干湿交替灌溉与植物生长调节剂对水稻光合特性及内源激素的影响[J]. 作物学报, 2025, 51(1): 174-188. |
| [2] | 钱玉平, 宿兵兵, 高吉星, 阮粉花, 李亚伟, 茅林春. 玉米大豆间作对喀斯特区土壤理化性质及微生物碳代谢特征的影响[J]. 作物学报, 2025, 51(1): 273-284. |
| [3] | 王丽萍, 李盼, 赵连豪, 樊志龙, 胡发龙, 范虹, 何蔚, 柴强, 殷文. 西北绿洲灌区玉米叶片衰老特征对不同地膜覆盖利用方式的响应[J]. 作物学报, 2025, 51(1): 233-246. |
| [4] | 王媛, 许佳茵, 董二伟, 王劲松, 刘秋霞, 黄晓磊, 焦晓燕. 有机肥替代化肥氮对谷子氮素累积、产量及品质的影响[J]. 作物学报, 2025, 51(1): 149-160. |
| [5] | 辛明华, 秘雅迪, 王国平, 李小飞, 李亚兵, 董合林, 韩迎春, 冯璐. 行距配置和种植密度对棉花干物质生产及产量的影响[J]. 作物学报, 2025, 51(1): 221-232. |
| [6] | 李超, 付小琼. 基于GYT双标图综合评价黄河流域中熟杂交棉花区域试验品种[J]. 作物学报, 2025, 51(1): 30-43. |
| [7] | 丁树启, 程彤, 王弼琨, 于德彬, 饶德民, 孟凡钢, 赵胤凯, 王晓慧, 张伟. 密植对不同年代大豆品种群体光合生产和产量形成的影响[J]. 作物学报, 2025, 51(1): 161-173. |
| [8] | 张军, 胡川, 周起晖, 任开明, 董誓言, 刘傲寒, 吴金芝, 黄明, 李友军. 减氮及有机肥替代对旱地冬小麦干物质积累、转运、分配和产量的影响[J]. 作物学报, 2025, 51(1): 207-220. |
| [9] | 张贵芹, 王洪章, 郭新送, 朱福军, 高涵, 张吉旺, 赵斌, 任佰朝, 刘鹏, 任昊. 有机物料投入对滨海盐碱地土壤理化性状和夏玉米产量形成的影响[J]. 作物学报, 2024, 50(9): 2323-2334. |
| [10] | 张琪祺, 陈杰昌, 蒯婕, 汪波, 王晶, 徐正华, 赵杰, 赵思明, 贾才华, 周广生. 高密度直播对油菜冷榨菜籽油品质的影响[J]. 作物学报, 2024, 50(9): 2358-2370. |
| [11] | 张振, 何建宁, 石玉, 于振文, 张永丽. 行距和种植方式对小麦光合特性和产量的影响[J]. 作物学报, 2024, 50(9): 2396-2407. |
| [12] | 徐一帆, 徐彩龙, 李瑞东, 吴宗声, 华建鑫, 杨琳, 宋雯雯, 吴存祥. 侧深施肥通过优化叶片功能与氮素积累来提高大豆产量[J]. 作物学报, 2024, 50(9): 2335-2346. |
| [13] | 杨煜琛, 靳雅荣, 骆金婵, 祝鑫, 李葳航, 贾纪原, 王小珊, 黄德均, 黄琳凯. 珍珠粟WD40基因家族鉴定及表达特征分析[J]. 作物学报, 2024, 50(9): 2219-2236. |
| [14] | 刘志鹏, 苟志文, 柴强, 殷文, 樊志龙, 胡发龙, 范虹, 王琦明. 干旱灌区绿肥对多样化种植小麦玉米产量性能指标的影响[J]. 作物学报, 2024, 50(9): 2415-2424. |
| [15] | 孙照华, 任昊, 王洪章, 王子强, 姚海燕, 辛爱美, 赵斌, 张吉旺, 任佰朝, 刘鹏. 叶面喷施硅制剂对滨海盐碱地夏玉米叶片光合性能及籽粒产量的影响[J]. 作物学报, 2024, 50(9): 2383-2395. |
|