欢迎访问作物学报,今天是
作物学报 ›› 2025, Vol. 51 ›› Issue (8): 2020-2032.doi: 10.3724/SP.J.1006.2025.51020
蔡金珊1( ), 李超男2, 王景一2, 李宁1, 柳玉平2, 景蕊莲2, 李龙2,*(), 孙黛珍1,*()
), 李超男2, 王景一2, 李宁1, 柳玉平2, 景蕊莲2, 李龙2,*(), 孙黛珍1,*()
CAI Jin-Shan1(), LI Chao-Nan2, WANG Jing-Yi2, LI Ning1, LIU Yu-Ping2, JING Rui-Lian2, LI Long2,*(), SUN Dai-Zhen1,*()
摘要:
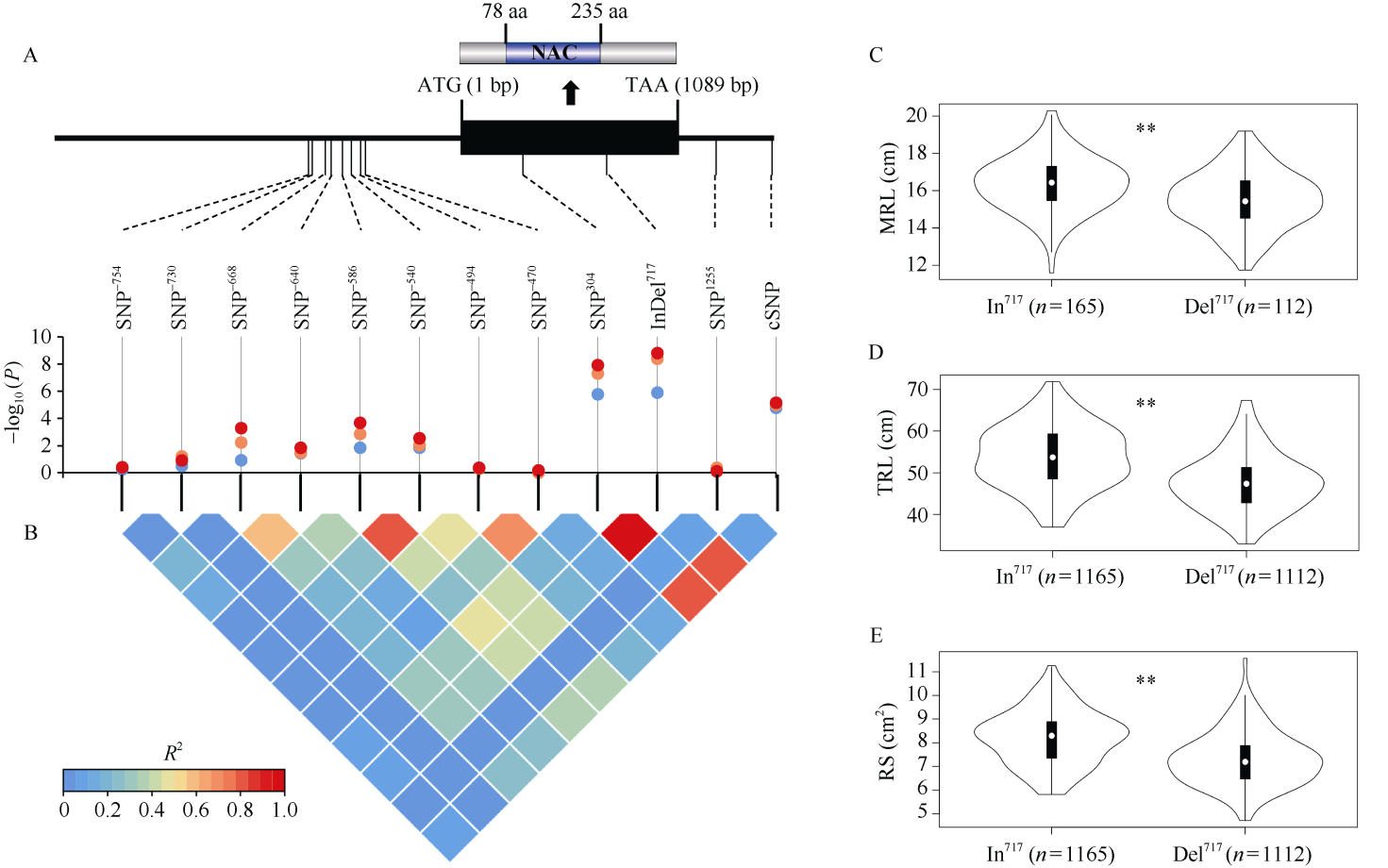
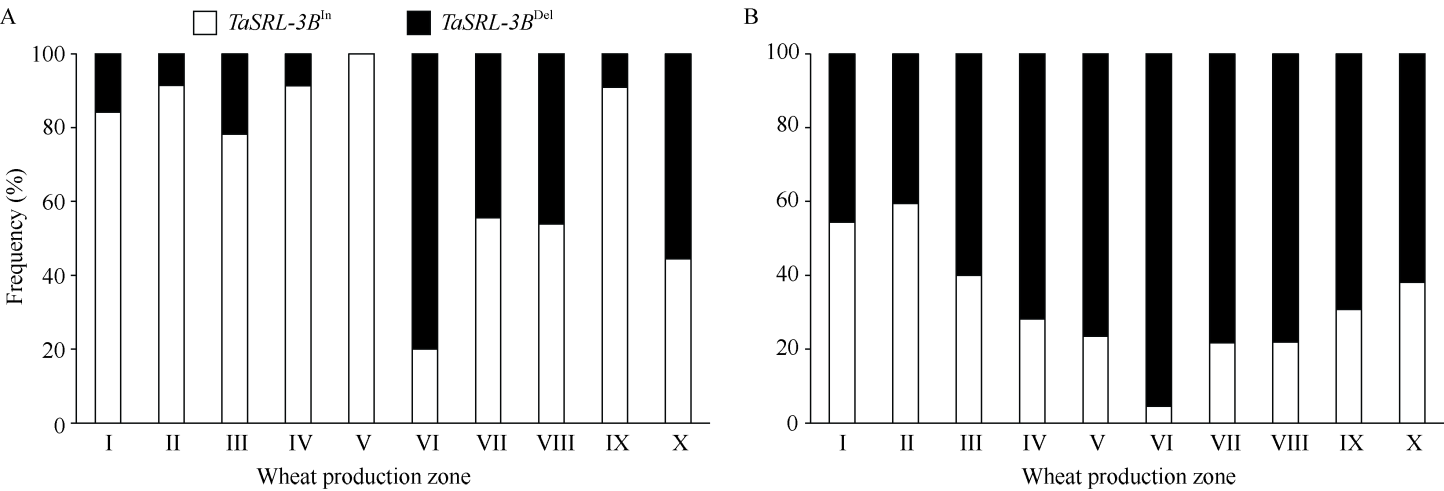
根系是小麦吸收土壤水分和养分的器官, 其形态特征与产量及耐逆性密切相关。因此, 发掘根系形态相关遗传位点及优异等位基因对于小麦改良具有重要意义。本项目以277份小麦种质为材料, 采用凝胶根室法鉴定总根长、根表面积及根角度等8种幼苗根系性状, 结合小麦660K SNP芯片的分型结果开展3种模型(GLM、MLM和FarmCPU)全基因组关联分析(GWAS)。共检测到52个关联位点, 其中包括6个与多个根系性状相关的一因多效性遗传位点(Loci17、Loci20、Loci22、Loci38、Loci46、Loci47), 分别位于染色体3A、3B、3D、5A、6A和6B上。在位点Loci20中克隆到调控根系性状候选基因TaSRL-3B, 其序列全长1089 bp, 无内含子, 第78~235位氨基酸处有1个保守的NAC结构域。在该基因编码区检测到1个20 bp的插入/缺失变异(InDel717), 该变异导致移码突变且与Loci20位点的候选SNP (cSNP, AX-108758584)紧密连锁(R2 = 0.84)。277份供试小麦材料中携带等位基因TaSRL-3BIn的种质平均最大根长、总根长及根表面积均显著大于携带等位基因TaSRL-3BDel的种质。以携带TaSRL-3BDel的鲁麦14 (LM14)为受体亲本、携带TaSRL-3BIn的陕合6号 (SH6)为供体亲本, 创制回交导入系群体(BC3F5)。利用基于InDel717开发的分子标记从中鉴定出5个携带TaSRL-3BIn的鲁麦14近等基因系。与鲁麦14相比, 其近等基因系的最大根长、总根长、根表面积及根体积均显著增加, 进一步表明TaSRL-3B参与调控小麦幼苗根系形态。与小麦地方品种相比, 我国现代育成品种中长根型等位基因TaSRL-3BIn频率减少。本研究为加快小麦根系遗传调控网络构建和功能解析提供了重要信息, 有助于小麦根系的遗传改良。
| [1] | FAOSTAT. Statistics Database.Rome Available at: http://www.fao.org/statistics/databases/en/ [2025-04-04]. |
| [2] | Bailey-Serres J, Parker J E, Ainsworth E A, Oldroyd G E D, Schroeder J I. Genetic strategies for improving crop yields. Nature, 2019, 575: 109-118. |
| [3] | Li L, Wang J Y, Li C N, Mao X G, Zhang X Q, Sun J W, Zhang K, Liu Y P, Reynolds M P, Yang Z G, et al. Insights into progress of wheat breeding in arid and infertile areas of China in the last 14 years. Field Crops Res, 2024, 306: 109220. |
| [4] | Nirmalaruban R, Yadav R, Sugumar S, Meda A, Babu P, Kumar M, Gaikwad K B, Bainsla N K, Singh S K, Suvitha R, et al. Root traits: a key for breeding climate-smart wheat (Triticum aestivum). Plant Breed, 2025, 144: 310-334. |
| [5] |
Ober E S, Alahmad S, Cockram J, Forestan C, Hickey L T, Kant J, Maccaferri M, Marr E, Milner M, Pinto F, et al. Wheat root systems as a breeding target for climate resilience. Theor Appl Genet, 2021, 134: 1645-1662.
doi: 10.1007/s00122-021-03819-w pmid: 33900415 |
| [6] | Tiwari V K, Saripalli G, Sharma P K, Poland J. Wheat genomics: genomes, pangenomes, and beyond. Trends Genet, 2024, 40: 982-992. |
| [7] | Sahito J H, Zhang H, Gishkori Z G N, Ma C H, Wang Z H, Ding D, Zhang X H, Tang J H. Advancements and prospects of genome-wide association studies (GWAS) in maize. Int J Mol Sci, 2024, 25: 1918. |
| [8] |
Han S C, Wang Y L, Li Y X, Zhu R, Gu Y S, Li J, Guo H F, Ye W, Nabi H G, Yang T, et al. The OsNAC41-RoLe1-OsAGAP module promotes root development and drought resistance in upland rice. Mol Plant, 2024, 17: 1573-1593.
doi: 10.1016/j.molp.2024.09.002 pmid: 39228126 |
| [9] | Li C H, Guo J, Wang D M, Chen X J, Guan H H, Li Y X, Zhang D F, Liu X Y, He G H, Wang T Y, et al. Genomic insight into changes of root architecture under drought stress in maize. Plant Cell Environ, 2023, 46: 1860-1872. |
| [10] | International Wheat Genome Sequencing Consortium (IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science, 2018, 361: eaar7191. |
| [11] | Yao Y Y, Guo W L, Gou J Y, Hu Z R, Liu J, Ma J, Zong Y, Xin M M, Chen W, Li Q, et al. Wheat2035: integrating pan-omics and advanced biotechnology for future wheat design. Mol Plant, 2025, 18: 272-297. |
| [12] | Jiao C Z, Xie X M, Hao C Y, Chen L Y, Xie Y X, Garg V, Zhao L, Wang Z H, Zhang Y Q, Li T, et al. Pan-genome bridges wheat structural variations with habitat and breeding. Nature, 2025, 637: 384-393. |
| [13] | Chen D D, Richardson T, Chai S C, Lynne McIntyre C, Rae A L, Xue G P. Drought-up-regulated TaNAC69-1 is a transcriptional repressor of TaSHY2 and TaIAA7, and enhances root length and biomass in wheat. Plant Cell Physiol, 2016, 57: 2076-2090. |
| [14] |
Wang D Z, Zhang X X, Cao Y, Batool A, Xu Y X, Qiao Y Z, Li Y P, Wang H, Lin X L, Bie X M, et al. TabHLH27 orchestrates root growth and drought tolerance to enhance water use efficiency in wheat. J Integr Plant Biol, 2024, 66: 1295-1312.
doi: 10.1111/jipb.13670 |
| [15] | Li Y Y, Zhang Y F, Li C N, Chen X, Yang L L, Zhang J, Wang J Y, Li L, Reynolds M P, Jing R L, et al. Transcription factor TaWRKY51 is a positive regulator in root architecture and grain yield contributing traits. Front Plant Sci, 2021, 12: 734614. |
| [16] | Yang W, Feng M, Yu K H, Cao J, Cui G X, Zhang Y M, Peng H R, Yao Y Y, Hu Z R, Ni Z F, et al. The TaCLE24b peptide signaling cascade modulates lateral root development and drought tolerance in wheat. Nat Commun, 2025, 16: 1952. |
| [17] | Li L, Mao X G, Wang J Y, Chang X P, Reynolds M, Jing R L. Genetic dissection of drought and heat-responsive agronomic traits in wheat. Plant Cell Environ, 2019, 42: 2540-2553. |
| [18] |
赵阳, 李龙, 杨进文, 景蕊莲, 孙黛珍, 王景一. 小麦E3泛素连接酶基因TaSINA-3A与多种环境下的株高和千粒重相关. 作物学报, 2024, 50: 2654-2664.
doi: 10.3724/SP.J.1006.2024.41008 |
|
Zhao Y, Li L, Yang J W, Jing R L, Sun D Z, Wang J Y. An E3 ubiquitin ligase gene TaSINA-3A is associated with plant height and 1000-grain weight in various environments in wheat. Acta Agron Sin, 2024, 50: 2654-2664 (in Chinese with English abstract).
doi: 10.3724/SP.J.1006.2024.41008 |
|
| [19] | Yin L L, Zhang H H, Tang Z S, Xu J Y, Yin D, Zhang Z W, Yuan X H, Zhu M J, Zhao S H, Li X Y, et al. rMVP: a memory- efficient, visualization-enhanced, and parallel-accelerated tool for genome-wide association study. Genom Proteom Bioinform, 2021, 19: 619-628. |
| [20] | Li M X, Yeung J M Y, Cherny S S, Sham P C. Evaluating the effective numbers of independent tests and significant p-value thresholds in commercial genotyping arrays and public imputation reference datasets. Hum Genet, 2012, 131: 747-756. |
| [21] | Chen Y M, Guo Y W, Guan P F, Wang Y F, Wang X B, Wang Z H, Qin Z, Ma S W, Xin M M, Hu Z R, et al. A wheat integrative regulatory network from large-scale complementary functional datasets enables trait-associated gene discovery for crop improvement. Mol Plant, 2023, 16: 393-414. |
| [22] |
Li L, Peng Z, Mao X G, Wang J Y, Li C N, Chang X P, Jing R L. Genetic insights into natural variation underlying salt tolerance in wheat. J Exp Bot, 2021, 72: 1135-1150.
doi: 10.1093/jxb/eraa500 pmid: 33130904 |
| [23] |
李龙, 李超男, 毛新国, 王景一, 景蕊莲. 作物根系表型鉴定评价方法的现状与展望. 中国农业科学, 2022, 55: 425-437.
doi: 10.3864/j.issn.0578-1752.2022.03.001 |
|
Li L, Li C N, Mao X G, Wang J Y, Jing R L. Advances and perspectives of approaches to phenotyping crop root system. Sci Agric Sin, 2022, 55: 425-437 (in Chinese with English abstract).
doi: 10.3864/j.issn.0578-1752.2022.03.001 |
|
| [24] | Urfan M, Sharma S, Hakla H R, Rajput P, Andotra S, Lehana P K, Bhardwaj R, Khan M S, Das R, Kumar S, et al. Recent trends in root phenomics of plant systems with available methods-discrepancies and consonances. Physiol Mol Biol Plants, 2022, 28: 1311-1321. |
| [25] |
Xiong H Y, He H D, Chang Y, Miao B B, Liu Z W, Wang Q Q, Dong F M, Xiong L Z. Multiple roles of NAC transcription factors in plant development and stress responses. J Integr Plant Biol, 2025, 67: 510-538.
doi: 10.1111/jipb.13854 |
| [26] | Xie C T, Li C L, Wang F X, Zhang F, Liu J J, Wang J X, Zhang X S, Kong X P, Ding Z J. NAC1 regulates root ground tissue maturation by coordinating with the SCR/SHR-CYCD6;1 module in Arabidopsis. Mol Plant, 2023, 16: 709-725. |
| [27] | Xu P P, Ma W, Hu J B, Cai W M. The nitrate-inducible NAC transcription factor NAC056 controls nitrate assimilation and promotes lateral root growth in Arabidopsis thaliana. PLoS Genet, 2022, 18: e1010090. |
| [28] |
Mao C J, He J M, Liu L N, Deng Q M, Yao X F, Liu C M, Qiao Y L, Li P, Ming F. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development. Plant Biotechnol J, 2020, 18: 429-442.
doi: 10.1111/pbi.13209 pmid: 31389120 |
| [29] | Swain N, Sahoo R K, Jeughale K P, Sarkar S, Selvaraj S, Parameswaran C, Katara J, Bose L K, Samantaray S. Rice homolog of Arabidopsis Xylem NAC domain 1 (OsXND1), a NAC transcription factor regulates drought stress responsive root system architecture in indica rice. Mol Genet Genomics, 2024, 299: 94. |
| [30] | Lyu S K, Guo H, Zhang M, Wang Q H, Zhang H, Ji W Q. Large-scale cloning and comparative analysis of TaNAC genes in response to stripe rust and powdery mildew in wheat (Triticum aestivum L.). Genes, 2020, 11: 1073. |
| [31] | Gao J, Zhao Y, Zhao Z K, Liu W, Jiang C H, Li J J, Zhang Z Y, Zhang H L, Zhang Y G, Wang X N, et al. RRS1 shapes robust root system to enhance drought resistance in rice. New Phytol, 2023, 238: 1146-1162. |
| [32] | Uga Y. Challenges to design-oriented breeding of root system architecture adapted to climate change. Breed Sci, 2021, 71: 3-12. |
| [33] | Voss-Fels K P, Qian L W, Parra-Londono S, Uptmoor R, Frisch M, Keeble-Gagnère G, Appels R, Snowdon R J. Linkage drag constrains the roots of modern wheat. Plant Cell Environ, 2017, 40: 717-725. |
| [34] | Xiang Y H, Yu J J, Liao B, Shan J X, Ye W W, Dong N Q, Guo T, Kan Y, Zhang H, Yang Y B, et al. An α/β hydrolase family member negatively regulates salt tolerance but promotes flowering through three distinct functions in rice. Mol Plant, 2022, 15: 1908-1930. |
| [1] | 杨婷婷, 陈娟, ABDUL Rehman, 李婧, 闫素辉, 汪建来, 李文阳. 花后弱光对软质小麦干物质积累转运、籽粒产量和淀粉品质的影响[J]. 作物学报, 2025, 51(8): 2204-2219. |
| [2] | 张飞飞, 何万龙, 焦文娟, 白斌, 耿洪伟, 程宇坤. 小麦抗条锈病相关性状元分析及候选基因分析[J]. 作物学报, 2025, 51(8): 2111-2127. |
| [3] | 闫喆林, 任强, 樊志龙, 殷文, 孙亚丽, 范虹, 何蔚, 胡发龙, 闫丽娟, 柴强. 氮肥后移优化绿洲灌区小麦间作玉米种间关系提高氮素利用效率[J]. 作物学报, 2025, 51(8): 2190-2203. |
| [4] | 宋改利, 王璐倩, 屈柯飞, 唐建卫, 董纯豪, 黄振朴, 高艳, 牛吉山, 殷贵鸿, 李巧云. Bipolaris sorokiniana黑胚病对中筋小麦淀粉含量、粒度分布与糊化特性的影响[J]. 作物学报, 2025, 51(8): 2164-2175. |
| [5] | 王曜阔, 王文政, 张敏, 刘希伟, 杨敏, 李昊昱, 张灵鑫, 闫彦菲, 蔡瑞国. 水氮运筹对冬小麦籽粒GMP合成和面粉加工品质的影响[J]. 作物学报, 2025, 51(8): 2176-2189. |
| [6] | 李宜谦, 徐守振, 刘萍, 马麒, 谢斌, 陈红. 基于40K SNP芯片的陆地棉产量构成因素全基因组关联分析及单铃重位点挖掘[J]. 作物学报, 2025, 51(8): 2128-2138. |
| [7] | 高梦娟, 赵贺莹, 陈家辉, 陈晓倩, 牛萌康, 钱琪润, 崔陆飞, 邢江敏, 银庆淼, 郭雯, 张宁, 孙丛苇, 阳霞, 裴丹, 贾奥琳, 陈锋, 余晓东, 任妍. 小麦抗纹枯病新位点Qse.hnau-5AS的定位及其候选基因鉴定[J]. 作物学报, 2025, 51(8): 2240-2250. |
| [8] | 姜朋, 吴磊, 黄倩楠, 李畅, 王化敦, 何漪, 张鹏, 张旭. 矮秆基因Rht-D1在长江中下游麦区的育种利用探索[J]. 作物学报, 2025, 51(8): 2077-2086. |
| [9] | 鲁向前, 付玉洁, 赵俊恒, 郑楠楠, 孙楠楠, 张国平, 叶玲珍. 小麦花药培养最佳取样时期穗部形态特征鉴定与高培养力基因型筛选[J]. 作物学报, 2025, 51(8): 2033-2047. |
| [10] | 吴柳格, 陈坚, 张鑫, 邓艾兴, 宋振伟, 郑成岩, 张卫建. 近二十年国审冬小麦品种的产量与品质性状变化趋势研究[J]. 作物学报, 2025, 51(7): 1814-1826. |
| [11] | 赵佳雯, 李子洪, 欧星雨, 王伊朗, 丁小飞, 梁乐瑶, 丁文金, 张海鹏, 马尚宇, 樊永惠, 黄正来, 张文静. 氮肥与钾肥运筹对弱筋小麦籽粒产量、品质的影响[J]. 作物学报, 2025, 51(7): 1914-1933. |
| [12] | 赵超男, 王金凤, 张玉, 张丽, 李瑞琦, 王鹏飞, 李鸽子, 张宏军, 虞波, 康国章. 全基因组关联分析定位与挖掘小麦氮高效基因[J]. 作物学报, 2025, 51(7): 1801-1813. |
| [13] | 王天译, 杨绣娟, 赵佳佳, 郝宇琼, 郑兴卫, 武棒棒, 李晓华, 郝水源, 郑军. 山西小麦醇溶蛋白多样性及其对面粉品质效应研究[J]. 作物学报, 2025, 51(7): 1784-1800. |
| [14] | 胡蒙, 沙丹, 张晟瑞, 谷勇哲, 张世碧, 李静, 孙君明, 邱丽娟, 李斌. 大豆分枝数QTL定位及候选基因筛选[J]. 作物学报, 2025, 51(7): 1747-1756. |
| [15] | 陈如雪, 孙丽芳, 张芯源, 牟海萌, 张永新, 袁丽雪, 彭仕乐, 王壮壮, 王永华. 秸秆还田与微生物菌剂配施对冬小麦旗叶碳氮代谢及产量形成的影响[J]. 作物学报, 2025, 51(7): 1901-1913. |
|
||