欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (3): 472-480.doi: 10.3724/SP.J.1006.2021.03027
周练, 刘朝显, 熊雨涵, 周京, 蔡一林*( )
)
ZHOU Lian, LIU Chao-Xian, XIONG Yu-Han, ZHOU Jing, CAI Yi-Lin*()
摘要:
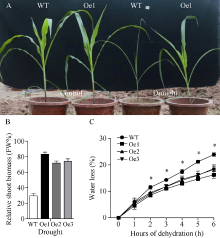
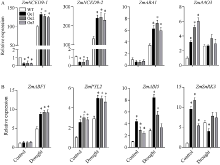
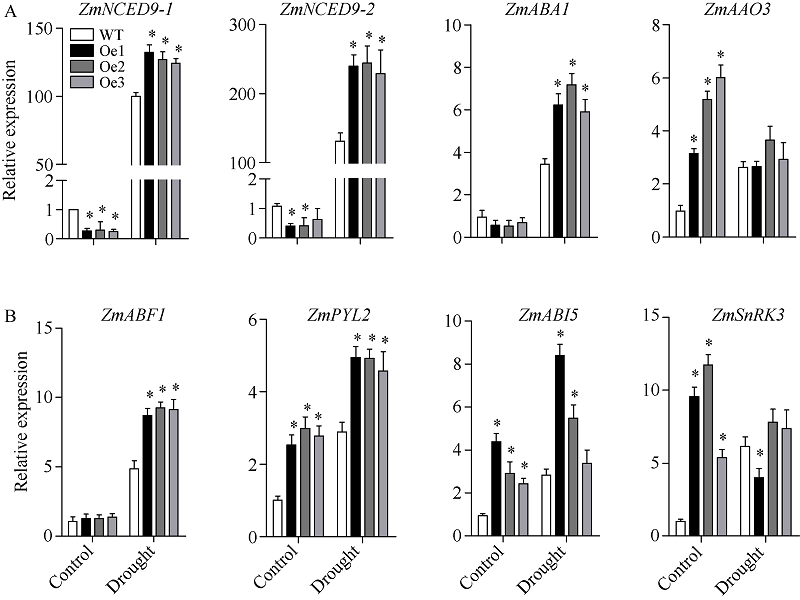
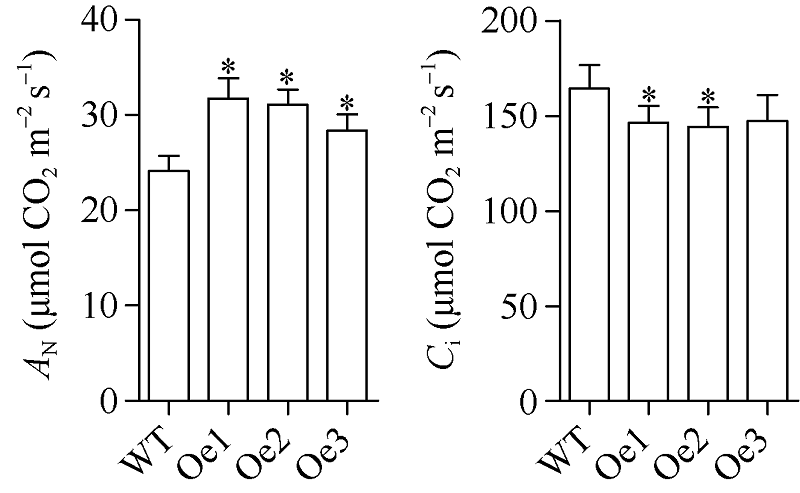
质膜内在蛋白(plasma membrane intrinsic proteins, PIPs)是水通道蛋白主要亚家族成员之一, 在植物生长发育过程中具有重要调节功能。前期研究表明, 玉米ZmPIP1;1基因表达受到渗透和盐胁迫的强烈诱导, 但其在玉米中的生物学功能尚不明确。本研究通过玉米遗传转化获得了ZmPIP1;1超表达转基因株系, 干旱胁迫实验揭示了ZmPIP1;1超表达转基因株系较野生型具有较低的水分散失率及较强的干旱胁迫耐性。转录组测序结果表明参与ABA生物合成及其信号通路相关基因的表达水平发生了显著变化。在田间正常生长条件下, ZmPIP1;1超表达转基因植株与野生型在生长发育过程中没有明显差异, 但转基因玉米株系具有较高的光合效率, 粒宽和百粒重增加, 玉米单果穗的产量提高。此外, 通过荧光双分子互补实验观察到ZmPIP1;1和ZmPIP2;6蛋白在玉米叶肉细胞原生质体的细胞质膜和叶绿体膜上存在互作, 并且可能导致了ZmPIP蛋白的重定位。该研究为ZmPIP1;1分子机制的解析奠定了重要基础, 为玉米高光效分子设计育种开辟了新的途径。
| [1] |
Yu C. China’s water crisis needs more than words. Nature, 2011,470:307.
pmid: 21331001 |
| [2] |
Long S P, Zhu X G, Naidu S L, Ort D R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ, 2006,29:315-330.
doi: 10.1111/j.1365-3040.2005.01493.x pmid: 17080588 |
| [3] | Nowicka B, Ciura J, Szymanska R, Kruk J. Improving photosynthesis, plant productivity and abiotic stress tolerance- current trends and future perspectives. J Plant Physiol, 2018,231:415-433. |
| [4] | Chaumont F, Moshelion M, Daniels M J. Regulation of plant aquaporin activity. Biol Cell, 2005,97:749-764. |
| [5] |
Maurel C. Plant aquaporins: novel functions and regulation properties. FEBS Lett, 2007,581:2227-2236.
pmid: 17382935 |
| [6] |
Kaldenhoff R, Ribas-Carbo M, Sans J F, Lovisolo C, Heckwolf M, Uehlein N. Aquaporins and plant water balance. Plant Cell Environ, 2008,31:658-666.
pmid: 18266903 |
| [7] |
Chaumont F, Tyerman S D. Aquaporins: highly regulated channels controlling plant water relations. Plant Physiol, 2014,164:1600-1618.
pmid: 24449709 |
| [8] | Chaumont F, Barrieu F, Wojcik E, Chrispeels M J, Jung R. Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol, 2001,125:1206-1215. |
| [9] |
Johanson U, Karlsson M, Johansson I, Gustavsson S, Sjovall S, Fraysse L, Weig A R, Kjellbom P. The complete set of genes encoding major intrinsic proteins inArabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiol, 2001,126:1358-1369.
doi: 10.1104/pp.126.4.1358 pmid: 11500536 |
| [10] | Ishibashi K. Aquaporin superfamily with unusual npa boxes: S-aquaporins (superfamily, sip-like and subcellular-aquaporins). Cell Mol Biol (Noisy-le-grand), 2006,52:20-27. |
| [11] |
Maurel C, Verdoucq L, Luu D T, Santoni V. Plant aquaporins: membrane channels with multiple integrated functions. Annu Rev Plant Biol, 2008,59:595-624.
pmid: 18444909 |
| [12] |
Danielson J A, Johanson U. Unexpected complexity of the aquaporin gene family in the moss Physcomitrella patens. BMC Plant Biol, 2008,8:45-59.
doi: 10.1186/1471-2229-8-45 pmid: 18430224 |
| [13] |
Bienert G P, Bienert M D, Jahn T P, Boutry M, Chaumont F. Solanaceae XIPs are plasma membrane aquaporins that facilitate the transport of many uncharged substrates. Plant J, 2011,66:306-317.
pmid: 21241387 |
| [14] |
Kammerloher W, Fischer U, Piechottka G P, Schaffner A R. Water channels in the plant plasma membrane cloned by immunoselection from a mammalian expression system. Plant J, 1994,6:187-199.
doi: 10.1046/j.1365-313x.1994.6020187.x pmid: 7920711 |
| [15] | Murata K, Mitsuoka K, Hirai T, Walz T, Agre P, Heymann J B, Engel A, Fujiyoshi Y. Structural determinants of water permeation through aquaporin-1. Nature, 2000,407:599-605. |
| [16] |
Fetter K, Van Wilder V, Moshelion M, Chaumont F. Interactions between plasma membrane aquaporins modulate their water channel activity. Plant Cell, 2004,16:215-228.
pmid: 14671024 |
| [17] | Zelazny E, Borst J W, Muylaert M, Batoko H, Hemminga M A, Chaumont F. FRET imaging in living maize cells reveals that plasma membrane aquaporins interact to regulate their subcellular localization. Proc Natl Acad Sci USA, 2007,104:12359-12364. |
| [18] |
Vandeleur R K, Mayo G, Shelden M C, Gilliham M, Kaiser B N, Tyerman S D. The role of plasma membrane intrinsic protein aquaporins in water transport through roots: diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine. Plant Physiol, 2009,149:445-460.
pmid: 18987216 |
| [19] | Chen W, Yin X, Wang L, Tian J, Yang R, Liu D, Yu Z, Ma N, Gao J. Involvement of rose aquaporin RhPIP1;1 in ethylene-regulated petal expansion through interaction with RhPIP2;1. Plant Mol Biol, 2013,83:219-233. |
| [20] | Yaneff A, Sigaut L, Marquez M, Alleva K, Pietrasanta L I, Amodeo G. Heteromerization of PIP aquaporins affects their intrinsic permeability. Proc Natl Acad Sci USA, 2014,111:231-236. |
| [21] | Afzal Z, Howton T C, Sun Y, Mukhtar M S. The roles of aquaporins in plant stress responses. J Dev Biol, 2016,4:9-30. |
| [22] | Alexandersson E, Danielson J A, Rade J, Moparthi V K, Fontes M, Kjellbom P, Johanson U. Transcriptional regulation of aquaporins in accessions ofArabidopsis in response to drought stress. Plant J, 2010,61:650-660. |
| [23] | Bae E K, Lee H, Lee J S, Noh E W. Drought, salt and wounding stress induce the expression of the plasma membrane intrinsic protein 1 gene in poplar (Populus alba × P. tremula var. glandulosa). Gene, 2011,483:43-48. |
| [24] | Peng Y, Lin W, Cai W, Arora R. Overexpression of aPanax ginseng tonoplast aquaporin alters salt tolerance, drought tolerance and cold acclimation ability in transgenic Arabidopsis plants. Planta, 2007,226:729-740. |
| [25] |
Zhou S, Hu W, Deng X, Ma Z, Chen L, Huang C, Wang C, Wang J, He Y, Yang G, He G. Overexpression of the wheat aquaporin gene,TaAQP7, enhances drought tolerance in transgenic tobacco. PLoS One, 2012,7:e52439.
pmid: 23285044 |
| [26] | Sreedharan S, Shekhawat U K, Ganapathi T R. Transgenic banana plants overexpressing a native plasma membrane aquaporin MusaPIP1;2 display high tolerance levels to different abiotic stresses. Plant Biotechnol J, 2013,11:942-952. |
| [27] | Zhou L, Zhou J, Xiong Y, Liu C, Wang J, Wang G, Cai Y. Overexpression of a maize plasma membrane intrinsic protein ZmPIP1;1 confers drought and salt tolerance in Arabidopsis. PLoS One, 2018,13:e0198639. |
| [28] | Uehlein N, Otto B, Hanson D T, Fischer M, McDowell N, Kaldenhoff R. Function ofNicotiana tabacum aquaporins as chloroplast gas pores challenges the concept of membrane CO2 permeability. Plant Cell, 2008,20:648-657. |
| [29] |
Hanba Y T, Shibasaka M, Hayashi Y, Hayakawa T, Kasamo K, Terashima I, Katsuhara M. Overexpression of the barley aquaporin HvPIP2;1 increases internal CO2 conductance and CO2 assimilation in the leaves of transgenic rice plants. Plant Cell Physiol, 2004,45:521-529.
doi: 10.1093/pcp/pch070 pmid: 15169933 |
| [30] |
Heckwolf M, Pater D, Hanson D T, Kaldenhoff R. TheArabidopsis thaliana aquaporin AtPIP1;2 is a physiologically relevant CO2 transport facilitator. Plant J, 2011,67:795-804.
pmid: 21564354 |
| [31] | 周练, 熊雨涵, 洪祥德, 周京, 刘朝显, 王久光, 王国强, 蔡一林. 玉米质膜内在蛋白ZmPIP2;6响应渗透、盐和干旱胁迫的功能鉴定. 中国农业科学, 53:461-473. |
| Zhou L, Xiong Y H, Hong X D, Zhou J, Liu C X, Wang J G, Wang G Q, Cai Y L. Functional characterization of a maize plasma membrane intrinsic protein ZmPIP2;6 responses to osmotic, salt and drought stress. Sci Agric Sin, 2020,53:461-473 (in Chinese with English abstract). | |
| [32] | Gookin T E, Assmann S M. Significant reduction of BiFC non-specific assembly facilitates in planta assessment of heterotrimeric G-protein interactors. Plant J, 2014,80:553-567. |
| [33] |
Nelson B K, Cai X, Nebenfuhr A. A multicolored set ofin vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J, 2007,51:1126-1136.
doi: 10.1111/j.1365-313X.2007.03212.x pmid: 17666025 |
| [34] | Bart R, Chern M, Park C J, Bartley L, Ronald P C. A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods, 2006,2:13-21. |
| [35] |
Nakashima K, Yamaguchi-Shinozaki K. ABA signaling in stress-response and seed development. Plant Cell Rep, 2013,32:959-970.
pmid: 23535869 |
| [36] |
Wang Z Y, Xiong L, Li W, Zhu J K, Zhu J. The plant cuticle is required for osmotic stress regulation of abscisic acid biosynthesis and osmotic stress tolerance in Arabidopsis. Plant Cell, 2011,23:1971-1984.
doi: 10.1105/tpc.110.081943 pmid: 21610183 |
| [37] | Lee S C, Lan W, Buchanan B B, Luan S. A protein kinase-phosphatase pair interacts with an ion channel to regulate ABA signaling in plant guard cells. Proc Natl Acad Sci USA, 2009,106:21419-21424. |
| [38] | Chater C, Peng K, Movahedi M, Dunn J A, Walker H J, Liang Y K, McLachlan D H, Casson S, Isner J C, Wilson I, Neill S J, Hedrich R, Gray J E, Hetherington A M. Elevated CO2-induced responses in stomata require ABA and ABA signaling. Curr Biol, 2015,25:2709-2716. |
| [39] | Sorrentino G, Haworth M, Wahbi S, Mahmood T, Zuomin S, Centritto M. Abscisic acid induces rapid reductions in mesophyll conductance to carbon dioxide. PLoS One, 2016,11:e0148554. |
| [40] |
Flexas J, Ribas-Carbo M, Hanson D T, Bota J, Otto B, Cifre J, McDowell N, Medrano H, Kaldenhoff R. Tobacco aquaporin NtAQP1 is involved in mesophyll conductance to CO2 in vivo. Plant J, 2006,48:427-439.
pmid: 17010114 |
| [41] | Ding L, Gao L, Liu W, Wang M, Gu M, Ren B, Xu G, Shen Q, Guo S. Aquaporin plays an important role in mediating chloroplastic CO2 concentration under high-N supply in rice (Oryza sativa) plants. Physiol Plant, 2016,156:215-226. |
| [42] |
Heinen R B, Bienert G P, Cohen D, Chevalier A S, Uehlein N, Hachez C, Kaldenhoff R, Le Thiec D, Chaumont F. Expression and characterization of plasma membrane aquaporins in stomatal complexes of Zea mays. Plant Mol Biol, 2014,86:335-350.
doi: 10.1007/s11103-014-0232-7 pmid: 25082269 |
| [43] |
Lin W, Peng Y, Li G, Arora R, Tang Z, Su W, Cai W. Isolation and functional characterization ofPgTIP1, a hormone-autotrophic cells-specific tonoplast aquaporin in ginseng. J Exp Bot, 2007,58:947-956.
pmid: 17237160 |
| [44] |
Zhou L, Wang C, Liu R, Han Q, Vandeleur R K, Du J, Tyerman S D, Shou H. Constitutive overexpression of soybean plasma membrane intrinsic protein GmPIP1;6 confers salt tolerance. BMC Plant Biol, 2014,14:181-193.
pmid: 24998596 |
| [45] | Patrick J W, Zhang W, Tyerman S D, Offler C E, Walker N A. Role of membrane transport in phloem translocation of assimilates and water. Funct Plant Biol, 2001,28:697-709. |
| [46] |
Liu F, Vantoai T, Moy L P, Bock G, Linford L D, Quackenbush J. Global transcription profiling reveals comprehensive insights into hypoxic response inArabidopsis. Plant Physiol, 2005,137:1115-1129.
doi: 10.1104/pp.104.055475 pmid: 15734912 |
| [47] |
Zhou Y, Setz N, Niemietz C, Qu H, Offler C E, Tyerman S D, Patrick J W. Aquaporins and unloading of phloem-imported water in coats of developing bean seeds. Plant Cell Environ, 2007,30:1566-1577.
doi: 10.1111/j.1365-3040.2007.01732.x pmid: 17927694 |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [4] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [5] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [6] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [7] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [8] | 王霞, 尹晓雨, 于晓明, 刘晓丹. 干旱锻炼对B73自交后代当代干旱胁迫记忆基因表达及其启动子区DNA甲基化的影响[J]. 作物学报, 2022, 48(5): 1191-1198. |
| [9] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [10] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [11] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [12] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [13] | 丁红, 徐扬, 张冠初, 秦斐斐, 戴良香, 张智猛. 不同生育期干旱与氮肥施用对花生氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 695-703. |
| [14] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [15] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
|
||