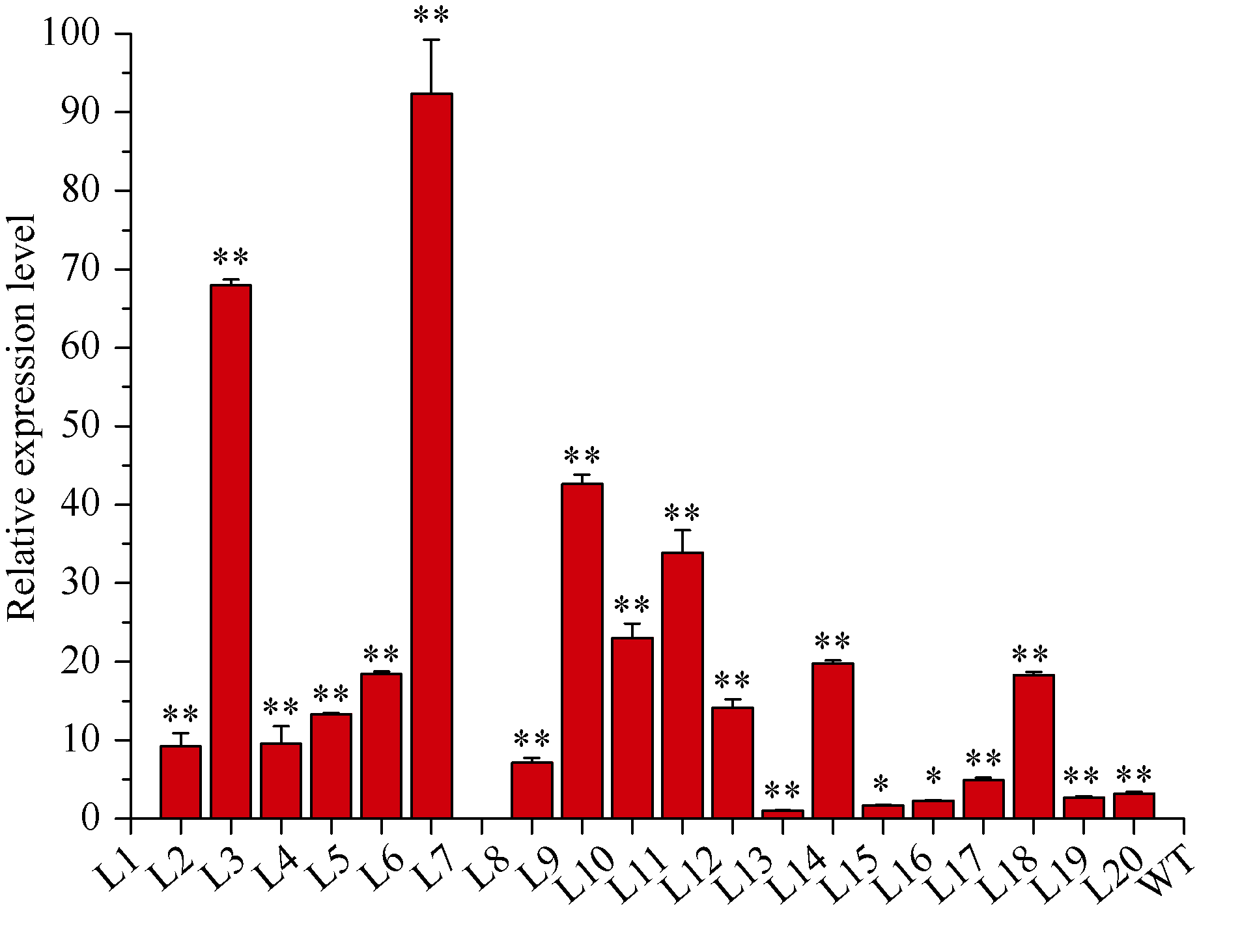
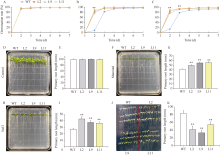
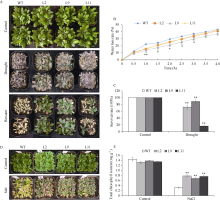
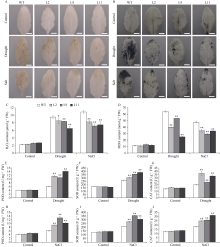
Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2025, Vol. 51 ›› Issue (3): 650-666.doi: 10.3724/SP.J.1006.2025.44082
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
HUO Ru-Xue1,2( ), GE Xiang-Han1, SHI Jia1, LI Xue-Rui1, DAI Sheng-Jie1, LIU Zhen-Ning1,*(), LI Zong-Yun2,*()
), GE Xiang-Han1, SHI Jia1, LI Xue-Rui1, DAI Sheng-Jie1, LIU Zhen-Ning1,*(), LI Zong-Yun2,*()
| [1] |
Mochida K, Yoshida T, Sakurai T, Yamaguchi-Shinozaki K, Shinozaki K, Tran L S. Genome-wide analysis of two-component systems and prediction of stress-responsive two-component system members in soybean. DNA Res, 2010, 17: 303-324.
doi: 10.1093/dnares/dsq021 pmid: 20817745 |
| [2] | Tran L S, Urao T, Qin F, Maruyama K, Kakimoto T, Shinozaki K, Yamaguchi-Shinozaki K. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc Natl Acad Sci USA, 2007, 104: 20623-20628. |
| [3] | Mason M G, Jha D, Salt D E, Tester M, Hill K, Kieber J J, Type-B response regulators ARR1 and ARR12 regulate expression of AtHKT1;1 and accumulation of sodium in Arabidopsis shoots. Plant J, 2010, 64: 753-763. |
| [4] |
Jain M, Tyagi A K, Khurana J P. Molecular characterization and differential expression of cytokinin-responsive type-a response regulators in rice (Oryza sativa). BMC Plant Biol, 2006, 6: 1.
pmid: 16472405 |
| [5] |
Le D T, Nishiyama R, Watanabe Y, Mochida K, Yamaguchi- Shinozaki K, Shinozaki K, Tran L S. Genome-wide expression profiling of soybean two-component system genes in soybean root and shoot tissues under dehydration stress. DNA Res, 2011, 18: 17-29.
doi: 10.1093/dnares/dsq032 pmid: 21208938 |
| [6] |
Jeon J, Kim J. Arabidopsis response regulator1 and Arabidopsis histidine phosphotransfer protein2 (AHP2), AHP3, and AHP5 function in cold signaling. Plant Physiol, 2013, 161: 408-424.
doi: 10.1104/pp.112.207621 pmid: 23124324 |
| [7] |
Nishiyama R, Watanabe Y, Leyva-Gonzalez M A, Ha C V, Fujita Y, Tanaka M, Seki M, Yamaguchi-Shinozaki K, Shinozaki K, Herrera-Estrella L, Tran L S P. Arabidopsis AHP2, AHP3, and AHP5 histidine phosphotransfer proteins function as redundant negative regulators of drought stress response. Proc Natl Acad Sci USA, 2013, 110: 4840-4845.
doi: 10.1073/pnas.1302265110 pmid: 23487796 |
| [8] | 潘雅姣, 王迪, 朱苓华, 傅彬英, 黎志康. 水稻双组分系统基因干旱胁迫表达谱分析. 作物学报, 2009, 35: 1628-1636. |
| Pan Y J, Wang D, Zhu L H, Fu B Y, Li Z K. Differential expressions of two-component element genes in rice under drought stress. Acta Agron Sin, 2009, 35: 1628-1636 (in Chinese with English abstract). | |
| [9] |
Jeon J, Kim N Y, Kim S, Kang N Y, Novák O, Ku S J, Cho C, Lee D J, Lee E J, Strnad M, Kim J. A subset of cytokinin two-component signaling system plays a role in cold temperature stress response in Arabidopsis. J Biol Chem, 2010, 285: 23371-23386.
doi: 10.1074/jbc.M109.096644 pmid: 20463025 |
| [10] | Karan R, Singla-Pareek S L, Pareek A. Histidine kinase and response regulator genes as they relate to salinity tolerance in rice. Funct Integr Genomics, 2009, 9: 411-417. |
| [11] | Schaller G E, Kieber J J, Shiu S H. Two-component signaling elements and histidyl-aspartyl phosphorelays. Arabidopsis Book, 2008, 6: e0112. |
| [12] | Pareek A, Singh A, Kumar M, Kushwaha H R, Lynn A M, Singla-Pareek S L. Whole-genome analysis of Oryza sativa reveals similar architecture of two-component signaling machinery with Arabidopsis. Plant Physiol, 2006, 142: 380-397. |
| [13] | Zameer R, Sadaqat M, Fatima K, Fiaz S, Rasul S, Zafar H, Qayyum A, Nashat N, Raza A, Shah A N, Batool R, Azeem F, Sun S M, Chung G. Two-component system genes in Sorghum bicolor: genome-wide identification and expression profiling in response to environmental stresses. Front Genet, 2021, 12: 794305. |
| [14] |
Chu Z X, Ma Q, Lin Y X, Tang X L, Zhou Y Q, Zhu S W, Fan J, Cheng B J. Genome-wide identification, classification, and analysis of two-component signal system genes in maize. Genet Mol Res, 2011, 10: 3316-3330.
doi: 10.4238/2011.December.8.3 pmid: 22194197 |
| [15] | Liu Z N, Zhang M, Kong L J, Lv Y X, Zou M H, Lu G, Cao J S, Yu X L. Genome-wide identification, phylogeny, duplication, and expression analyses of two-component system genes in Chinese cabbage (Brassica rapa ssp. pekinensis). DNA Res, 2014, 21: 379-396. |
| [16] | Gahlaut V, Mathur S, Dhariwal R, Khurana J P, Tyagi A K, Balyan H S, Gupta P K. A multi-step phosphorelay two-component system impacts on tolerance against dehydration stress in common wheat. Funct Integr Genomics, 2014, 14: 707-716. |
| [17] | He Y J, Liu X, Ye L, Pan C T, Chen L F, Zou T, Lu G. Genome- wide identification and expression analysis of two-component system genes in tomato. Int J Mol Sci, 2016, 17: 1204. |
| [18] | Ahmad B, Azeem F, Ali M A, Nawaz M A, Nadeem H, Abbas A, Batool R, Atif R M, Ijaz U, Nieves-Cordones M, Chung G. Genome-wide identification and expression analysis of two component system genes in Cicer arietinum. Genomics, 2020, 112: 1371-1383. |
| [19] | Huo R X, Liu Z N, Yu X L, Li Z Y. The interaction network and signaling specificity of two-component system in Arabidopsis. Int J Mol Sci, 2020, 21: 4898. |
| [20] |
Kakimoto T. CKI1, a histidine kinase homolog implicated in cytokinin signal transduction. Science, 1996, 274: 982-985.
pmid: 8875940 |
| [21] | Desikan R, Horák J, Chaban C, Mira-Rodado V, Witthöft J, Elgass K, Grefen C, Cheung M K, Meixner A J, Hooley R, Neill S J, Hancock J T, Harter K. The histidine kinase AHK5 integrates endogenous and environmental signals in Arabidopsis guard cells. PLoS One, 2008, 3: e2491. |
| [22] | Mira-Rodado V. New insights into multistep-phosphorelay (MSP)/two-component system (TCS) regulation: are plants and bacteria that different? Plants (Basel), 2019, 8: 590. |
| [23] | Iwama A, Yamashino T, Tanaka Y, Sakakibara H, Kakimoto T, Sato S, Kato T, Tabata S, Nagatani A, Mizuno T. AHK5 histidine kinase regulates root elongation through an ETR1-dependent abscisic acid and ethylene signaling pathway in Arabidopsis thaliana. Plant Cell Physiol, 2007, 48: 375-380. |
| [24] | Pham J, Liu J, Bennett M H, Mansfield J W, Desikan R. Arabidopsis histidine kinase 5 regulates salt sensitivity and resistance against bacterial and fungal infection. New Phytol, 2012, 194: 168-180. |
| [25] |
Pham J, Desikan R. Modulation of ROS production and hormone levels by AHK5 during abiotic and biotic stress signaling. Plant Signal Behav, 2012, 7: 893-897.
doi: 10.4161/psb.20692 pmid: 22827948 |
| [26] | Huo R X, Zhao Y S, Liu T X, Xu M, Wang X H, Xu P, Dai S J, Cui X Y, Han Y H, Liu Z N, Li Z Y. Genome-wide identification and expression analysis of two-component system genes in sweet potato (Ipomoea batatas L.). Front Plant Sci, 2023, 13: 1091620. |
| [27] | Yu Y C, Xuan Y, Bian X F, Zhang L, Pan Z Y, Kou M, Cao Q H, Tang Z H, Li Q, Ma D F, Li Z Y, Sun J. Overexpression of phosphatidylserine synthase IbPSS1 affords cellular Na+ homeostasis and salt tolerance by activating plasma membrane Na+/H+ antiport activity in sweet potato roots. Hortic Res, 2020, 7: 131. |
| [28] |
Clough S J, Bent A F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J, 1998, 16: 735-743.
doi: 10.1046/j.1365-313x.1998.00343.x pmid: 10069079 |
| [29] | Zhang X R, Henriques R, Lin S S, Niu Q W, Chua N H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat Protoc, 2006, 1: 641-646. |
| [30] | Yu H J, Hogan P, Sundaresan V. Analysis of the female gametophyte transcriptome of Arabidopsis by comparative expression profiling. Plant Physiol, 2005, 139: 1853-1869. |
| [31] | Park S C, Kim Y H, Ji C Y, Park S, Jeong J C, Lee H S, Kwak S S. Stable internal reference genes for the normalization of real-time PCR in different sweetpotato cultivars subjected to abiotic stress conditions. PLoS One, 2012, 7: e51502. |
| [32] | Szmitkowska A, Cuyacot A R, Pekaérovaé B, Žd'árská M, Houser J, Komárek J, Jaseňáková Z, Jayasree A, Heunemann M, Ubogoeva E, Spyroglou I, Trtílek M, Mironova V, Harter K, Zemlyanskaya E, Žídek L, Wimmerová M, Hejátko J. AHK5 mediates ETR1-initiated multistep phosphorelay in Arabidopsis. bioRxiv, 2021, DOI: 10.1101/2021.09.16.460643. |
| [33] |
He Y J, Liu X, Zou T, Pan C T, Qin L, Chen L F, Lu G. Genome-wide identification of two-component system genes in Cucurbitaceae crops and expression profiling analyses in cucumber. Front Plant Sci, 2016, 7: 899.
doi: 10.3389/fpls.2016.00899 pmid: 27446129 |
| [1] | WANG Yu-Xin, CHEN Tian-Yu, ZHAI Hong, ZHANG Huan, GAO Shao-Pei, HE Shao-Zhen, ZHAO Ning, LIU Qing-Chang. Cloning and characterization of drought tolerance function of kinase gene IbHT1 in sweetpotato [J]. Acta Agronomica Sinica, 2025, 51(2): 301-311. |
| [2] | MENG Fan-Hua, LIU Min, SHEN Ao, LIU Wei. Preliminary investigation of the SiLTP1: a lipid transfer protein gene involved in the salt tolerance of foxtail millet [J]. Acta Agronomica Sinica, 2025, 51(1): 58-67. |
| [3] | LIU Yong-Hui, SHEN Yi, SHEN Yue, LIANG Man, SHA Qin, ZHANG Xu-Yao, CHEN Zhi-De. Cloning and functional analysis of drought-inducible promoter AhMYB44-11- Pro in peanut (Arachis hypogaea L.) [J]. Acta Agronomica Sinica, 2024, 50(9): 2157-2166. |
| [4] | LI Wen-Juan, WANG Li-Min, QI Yan-Ni, ZHAO Wei, XIE Ya-Ping, DANG Zhao, ZHAO Li-Rong, LI Wen, XU Chen-Meng, WANG Yan, ZHANG Jian-Ping. Functional analysis of flax LuWRI1a in response to drought and salt stresses [J]. Acta Agronomica Sinica, 2024, 50(7): 1750-1761. |
| [5] | SHE Meng, ZHENG Deng-Yu, KE Zhao, WU Zhong-Yi, ZOU Hua-Wen, ZHANG Zhong-Bao. Cloning and functional analysis of ZmGRAS13 gene in maize [J]. Acta Agronomica Sinica, 2024, 50(6): 1420-1434. |
| [6] | WANG Long, LI Jing, QIAN Chen, LIN Guo-Bing, LI Yi-Yang, YANG Guang, ZUO Qing-Song. Effects of salt stress on yield, quality, and physiology in rapeseed [J]. Acta Agronomica Sinica, 2024, 50(6): 1597-1607. |
| [7] | QIAO Zhi-Xin, ZHANG Jie-Dao, WANG Yu, GUO Qi-Fang, LIU Yan-Jing, CHEN Rui, HU Wen-Hao, SUN Ai-Qing. Difference in germination characteristics of different winter wheat cultivars under drought stress [J]. Acta Agronomica Sinica, 2024, 50(6): 1568-1583. |
| [8] | ZHU Xiao-Ya, ZHANG Qiang-Qiang, ZHAO Peng, LIU Ming, WANG Jing, JIN Rong, YU Yong-Chao, TANG Zhong-Hou. Transcriptome and metabolomic analysis of foliar spraying of Salvia miltiorrhiza carbon dots to alleviate low phosphorus stress in sweetpotato [J]. Acta Agronomica Sinica, 2024, 50(2): 383-393. |
| [9] | ZHANG Jin-Hui, XIAO Zi-Yi, LI Xu-Hua, ZHANG Ming, JIA Chun-Lan, PAN Zhen-Yuan, QIU Fa-Zhan. Salt tolerance evaluation and transcriptome analysis of maize mutant caspl2b2 [J]. Acta Agronomica Sinica, 2024, 50(12): 3144-3154. |
| [10] | YANG Chuang, WANG Ling, QUAN Cheng-Tao, YU Liang-Qian, DAI Cheng, GUO Liang, FU Ting-Dong, MA Chao-Zhi. Relative expression profiles of genes response to salt stress and constructions of gene co-expression networks in Brassica napus L. [J]. Acta Agronomica Sinica, 2024, 50(1): 237-250. |
| [11] | XIAO Sheng-Hua, LU Yan, LI An-Zi, QIN Yao-Bin, LIAO Ming-Jing, BI Zhao-Fu, ZHUO Gan-Feng, ZHU Yong-Hong, ZHU Long-Fu. Function analysis of an AP2/ERF transcription factor GhTINY2 in cotton negatively regulating salt tolerance [J]. Acta Agronomica Sinica, 2024, 50(1): 126-137. |
| [12] | WANG Li-Ping, WANG Xiao-Yu, FU Jing-Ye, WANG Qiang. Functional identification of maize transcription factor ZmMYB12 to enhance drought resistance and low phosphorus tolerance in plants [J]. Acta Agronomica Sinica, 2024, 50(1): 76-88. |
| [13] | GUO Jia-Xin, YE Yang, GUO Hui-Juan, MIN Wei. Effects and variability analysis of different salt and alkali stresses on the proteome of cotton leaves [J]. Acta Agronomica Sinica, 2024, 50(1): 219-236. |
| [14] | XU Yang, ZHANG Dai, KANG Tao, WEN Sai-Qun, ZHANG Guan-Chu, DING Hong, GUO Qing, QIN Fei-Fei, DAI Liang-Xiang, ZHANG Zhi-Meng. Effects of salt stress on ion dynamics and the relative expression level of salt tolerance genes in peanut seedlings [J]. Acta Agronomica Sinica, 2023, 49(9): 2373-2384. |
| [15] | SU Yi-Jun, ZHAO Lu-Kuan, TANG Fen, DAI Xi-Bin, SUN Ya-Wei, ZHOU Zhi-Lin, LIU Ya-Ju, CAO Qing-He. Genetic diversity and population structure analysis of 378 introduced sweetpotato germplasm collections [J]. Acta Agronomica Sinica, 2023, 49(9): 2582-2593. |
|
||