欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (9): 2242-2254.doi: 10.3724/SP.J.1006.2022.11079
谭照国1,2( ), 苑少华2(), 李艳梅2, 白建芳2, 岳洁茹2, 刘子涵2, 张天豹2, 赵福永1, 赵昌平2, 许本波1, 张胜全2,*(), 庞斌双2,*(), 张立平1,2,*()
), 苑少华2(), 李艳梅2, 白建芳2, 岳洁茹2, 刘子涵2, 张天豹2, 赵福永1, 赵昌平2, 许本波1, 张胜全2,*(), 庞斌双2,*(), 张立平1,2,*()
TAN Zhao-Guo1,2(), YUAN Shao-Hua2(), LI Yan-Mei2, BAI Jian-Fang2, YUE Jie-Ru2, LIU Zi-Han2, ZHANG Tian-Bao2, ZHAO Fu-Yong1, ZHAO Chang-Ping2, XU Ben-Bo1, ZHANG Sheng-Quan2,*(), PANG Bin-Shuang2,*(), ZHNAG Li-Ping1,2,*()
摘要:
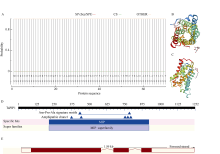
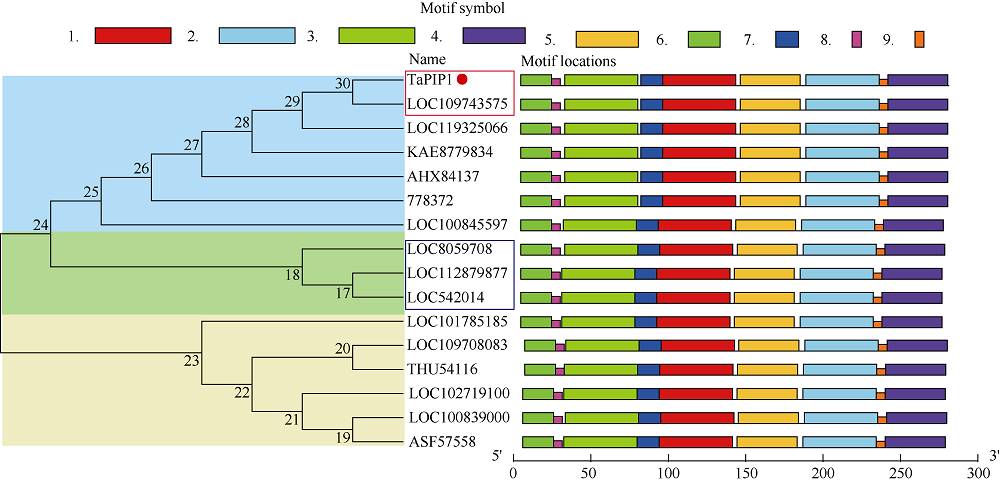
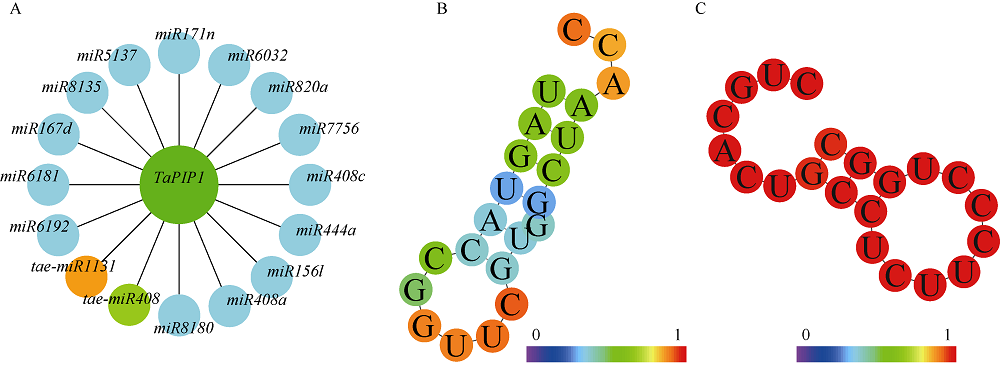
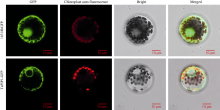
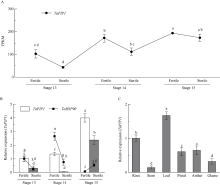
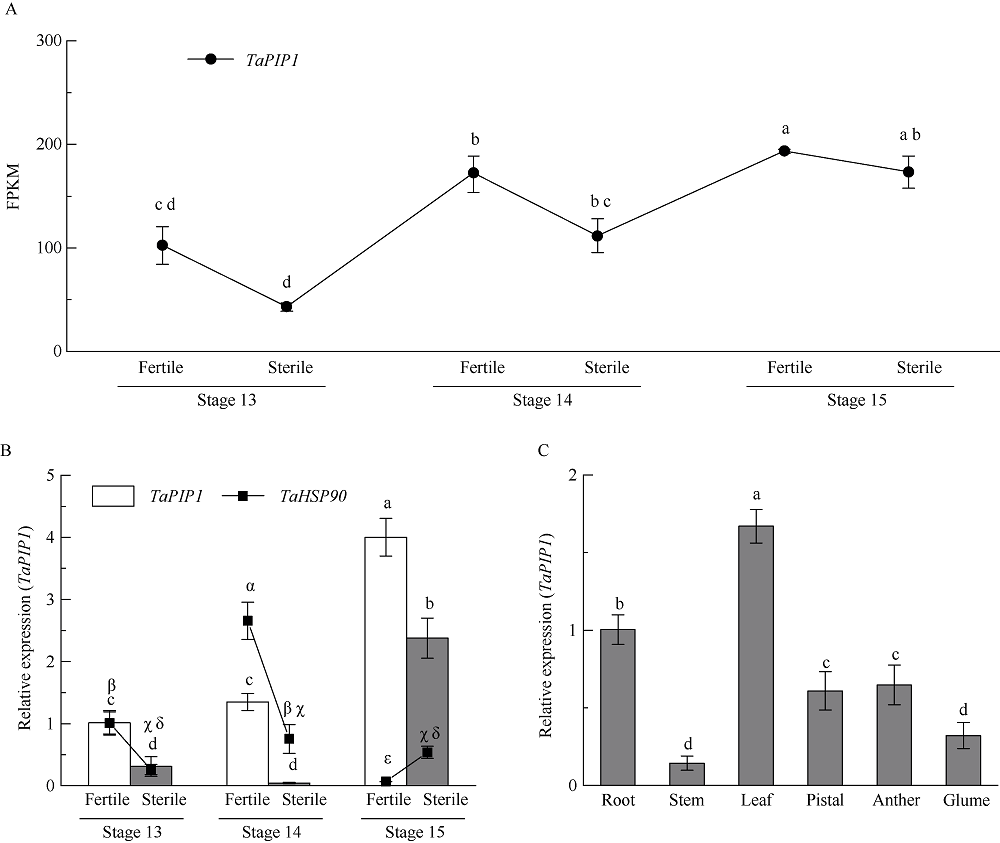
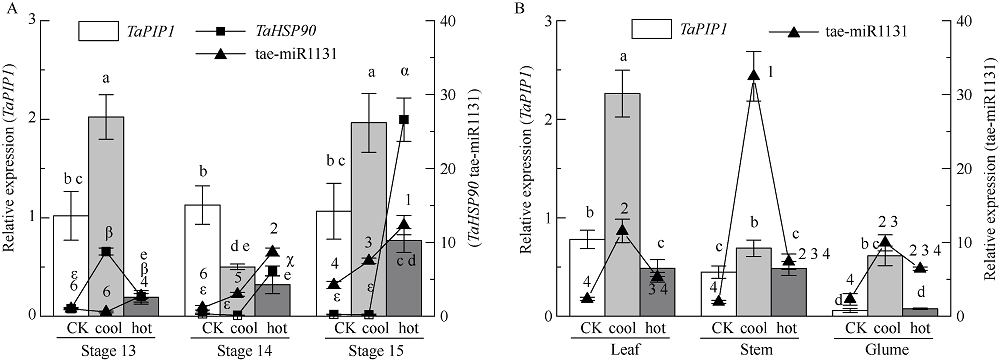
二系杂交小麦育种是小麦产量提高的重要途径之一。光温敏雄性不育小麦生殖生长过程中花药的发育及开裂情况直接影响杂交小麦的制种效率和产量。植物花药的开裂与脱水活动紧密相关。水通道蛋白(aquaporins, AQPs)是高效转运水分及特异小分子的膜内在蛋白, 其中质膜内在蛋白(plasma membrane intrinsic proteins, PIPs)在水分的吸收与外排中发挥着重要作用。为进一步了解水通道蛋白在小麦光温敏核雄性不育系花药开裂中的作用提供理论基础。本研究以不育系BS366花药的cDNA为模板, 克隆获得了TaPIP1基因。利用生物信息学软件对TaPIP1进行分析, 该基因包含一个879 bp的开放阅读框, 编码292个氨基酸。TaPIP1的启动子区存在赤霉素、脱落酸、茉莉酸和光等响应元件。TaPIP1属于MIP超家族, 具有典型的NPA保守结构域, 亚细胞定位于细胞质膜与核膜上。miRNA互作预测发现TaPIP1受tae-miR1131与tae-miR408的剪切抑制, 表明TaPIP1可能和与植物的抗氧化能力相关。通过蛋白互作预测及qPCR实验, 表明TaPIP1可与热激蛋白(heat shock protein 90, TaHSP90)相互作用, 在高温和干旱复合胁迫下, 参与花药细胞壁膨压调控, 进而调控花药的开裂。
| [1] |
Feng Z J, Xu S C, Liu N, Zhang G W, Hu Q Z, Xu Z S, Gong Y M. Identification of the AQP members involved in abiotic stress responses from Arabidopsis. Gene, 2018, 646: 64-73.
doi: 10.1016/j.gene.2017.12.048 |
| [2] | Chaumont F, Moshelion M, Daniels M J. Regulation of plant aquaporin activity. Biol Cell, 2005, 97: 749-764. |
| [3] |
Ishikawa F, Suga S, Uemura T, Sato M H, Maeshima M. Novel type aquaporin SIPs are mainly localized to the ER membrane and show cell-specific expression in Arabidopsis thaliana. FEBS Lett, 2005, 579: 5814-5820.
doi: 10.1016/j.febslet.2005.09.076 |
| [4] | Azad A K, Ahmed J, Alum M A, Hasan M M, Ishikawa T, Sawa Y, Katsuhara M. Genome-wide characterization of major intrinsic proteins in four grass plants and their non-aqua transport selectivity profiles with comparative perspective. PLoS One, 2016, 11: e0157735. |
| [5] |
Chaumont F, Barrieu F, Wojcik E, Chrispeels M J, Jung R. Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol, 2001, 125: 1206-1215.
pmid: 11244102 |
| [6] | Zhang D Y, Ali Z, Wang C B, Xu L, Yi J X, Xu Z L, Liu X Q, He X L, Huang Y H, Khan I A, Trethowan R M, Ma H X. Genome-wide sequence characterization and expression analysis of major intrinsic proteins in soybean (Glycine max L.). PLoS One, 2013, 8: e56312. |
| [7] |
Yuan D, Li W, Hua Y P, King G H J, Xu F S, Shi L. Genome-wide identification and characterization of the aquaporin gene family and transcriptional responses to boron deficiency in Brassica napus. Front Plant Sci, 2017, 8: 1336.
doi: 10.3389/fpls.2017.01336 pmid: 28824672 |
| [8] |
Javot H, Lauvergeat V, Santoni V, Martin-Laurent F, Guclu J, Vinh J, Heyes J, Franck K I, Schaffner A R, Bouchez D, Maurel C. Role of a single aquaporin isoform in root water uptake. Plant Cell, 2003, 15: 509-522.
doi: 10.1105/tpc.008888 |
| [9] |
Flexas J, Ribas-Carbo M, Hanson D T, Bota J, Otto B, Cifre J, McDowell N, Medrano H, Kaldenhoff R. Tobacco aquaporin NtAQP1 is involved in mesophyll conductance to CO2 in vivo. Plant J, 2006, 48: 427-439.
pmid: 17010114 |
| [10] |
Takano J, Wada M, Ludewig U, Schaaf G, von Wiren N, Fujiwara T. The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron uptake and plant development under boron limitation. Plant Cell, 2006, 18: 1498-1509.
doi: 10.1105/tpc.106.041640 |
| [11] |
Ma J F, Tamai K, Yamaji N, Mitani N, Konishi S, Katsuhara M, Ishiguro M, Murata Y, Yano M. A silicon transporter in rice. Nature, 2006, 440: 688-691.
doi: 10.1038/nature04590 |
| [12] |
Mao Z L, Sun W N. Arabidopsis seed-specific vacuolar aquaporins are involved in maintaining seed longevity under the control of ABSCISIC ACID INSENSITIVE 3. J Exp Bot, 2015, 66: 4781-4794.
doi: 10.1093/jxb/erv244 |
| [13] |
Secchi F, Schubert A, Lovisolo C. Changes in air CO2 concentration differentially alter transcript levels of NtAQP1 and NtPIP2;1 Aquaporin genes in tobacco leaves. Int J Mol Sci, 2016, 17: 567.
doi: 10.3390/ijms17040567 |
| [14] |
Uehlein N, Lovisolo C, Siefritz F, Kaldenhoff R. The tobacco aquaporin NtAQP1 is a membrane CO2 pore with physiological functions. Nature, 2003, 425: 734-737.
doi: 10.1038/nature02027 |
| [15] |
Holm L M, Jahn T P, Moller A L, Schjoerring J K, Ferri D, Klaerke D A, Zeuthen T. NH3 and NH4+ permeability in aquaporin-expressing Xenopus oocytes. Pflugers Arch-Eur J Physiol, 2005, 450: 415-428.
doi: 10.1007/s00424-005-1399-1 |
| [16] | Baluška F. Plant Aquaporins from Transport to Signaling. Signaling and Communication in Plants. Berlin: Springer Nature, 2017. pp 1-333. |
| [17] |
Azad A K, Sawa Y, Ishikawa T, Shibata H. Characterization of protein phosphatase 2A acting on phosphorylated plasma membrane aquaporin of tulip petals. Biosci Biotechnol Biochem, 2004, 68: 1170-1174.
doi: 10.1271/bbb.68.1170 |
| [18] |
Bai J F, Wang Y K, Wang P, Yuan S H, Gao J G, Duan W J, Wang N, Zhang F T, Zhang W J, Qin M Y, Zhao C P, Zhang L P. Genome-wide identification and analysis of the COI gene family in wheat (Triticum aestivum L.). BMC Genomics, 2018, 19: 754.
doi: 10.1186/s12864-018-5116-9 |
| [19] |
Wilson Z A, Song J, Taylor B, Yang C. The final split: the regulation of anther dehiscence. J Exp Bot, 2011, 62: 1633-1649.
doi: 10.1093/jxb/err014 pmid: 21325605 |
| [20] | Bots M, Feron R, Uehlein N, Weterings K, Kaldenhoff R, Mariani T. PIP1 and PIP2 aquaporins are differentially expressed during tobacco anther and stigma development. J Exp Bot, 2005, 56: 113-121. |
| [21] | 谭照国, 李艳梅, 白建芳, 郭昊宇, 栗婷婷, 段文静, 刘子涵, 苑少华, 张天豹, 张风廷, 陈兆波, 赵福永, 赵昌平, 张立平. 小麦TaBG的克隆及其在花药开裂中的潜在功能. 中国农业科学, 2021, 54: 2711-2723. |
| Tan Z G, Li Y M, Bai J F, Guo H Y, Li T T, Duan W J, Liu Z H, Yuan S H, Zhang T B, Zhang F T, Chen Z B, Zhao F Y, Zhao C P, Zhang L P. Cloning of TaBG and analysis of its function in anther dehiscence in wheat. Sci Agric Sin, 2021, 54: 2711-2723. (in Chinese with English abstract) | |
| [22] | 卢奕霏, 顾迎港, 陈威, 王娜, 康珍, 侯泽豪, 张迎新, 方正武, 马东方, 刘易科, 朱展望, 张改生, 王书平. 高温胁迫对小麦花药活性氧代谢的影响. 麦类作物学报, 2020, 40: 488-493. |
| Lu Y F, Gu Y G, Chen W, Wang N, Kang Z, Hou Z H, Zhang Y X, Fang Z W, Ma D F, Liu Y K, Zhu Z W, Zhang G S, Wang S P. Effect of high-temperature stress on reactive oxygen metabolism of wheat anther. J Triticeae Crops, 2020, 40: 488-493. (in Chinese with English abstract) | |
| [23] | 孙鹤, 郎志宏, 朱莉, 黄大昉. 玉米、小麦、水稻原生质体制备条件优化. 生物工程学报, 2013, 29: 224-234. |
| Sun H, Lang Z H, Zhu L, Huang D F. Optimized condition for protoplast isolation from maize, wheat and rice leaves. Chin J Biotech, 2013, 29: 224-234. (in Chinese with English abstract) | |
| [24] |
Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25: 402-408.
doi: 10.1006/meth.2001.1262 pmid: 11846609 |
| [25] | Tang J Y, Chu C C. MicroRNAs in crop improvement: fine-tuners for complex traits. Nature Plants, 2017, 3: 17077. |
| [26] |
Fotiadis D, Jeno P, Mini T, Wirtz S, Muller S A, Fraysse L, Kjellbom P, Engel A. Structural characterization of two aquaporins isolated from native spinach leaf plasma membranes. J Biol Chem, 2001, 276: 1707-1714.
doi: 10.1074/jbc.M009383200 pmid: 11050104 |
| [27] |
Hu J, Mitchum M G, Barnaby N, Ayele B T, Ogawa M, Nam E, Lai W C, Hanada A, Alonso J M, Ecker J R, Swain S M, Yamaguchi S, Kamiya Y, Sun T P. Potential sites of bioactive gibberellin production during reproductive growth in Arabidopsis Plant Cell, 2008, 20: 320-336.
doi: 10.1105/tpc.107.057752 |
| [28] |
Werner M, Uehlein N, Proksch P, Kaldenhoff R. Characterization of two tomato aquaporins and expression during the incompatible interaction of tomato with the plant parasite Cuscuta reflexa. Planta, 2001, 213: 550-555.
pmid: 11556787 |
| [29] |
Morillon R, Catterou M, Sangwan R S, Sangwan B S, Lassalles J P. Brassinolide may control aquaporin activities in Arabidopsis thaliana. Planta, 2001, 212: 199-204.
pmid: 11216840 |
| [30] |
Phillips A L, Huttly A K. Cloning of two gibberellin-regulated cDNAs from Arabidopsis thaliana by subtractive hybridization: expression of the tonoplast water channel, γ-TIP, is increased by GA3. Plant Mol Biol, 1994, 24: 603-615.
pmid: 8155880 |
| [31] |
Hu W, Yuan Q Q, Wang Y Y, Cai R, Deng X M, Wang J, Zhou S Y, Chen M J, Chen L H, Huang C, Ma Z B, Yang G X, He G Y. Overexpression of a wheat aquaporin gene, TaAQP8, enhances salt stress tolerance in transgenic tobacco. Plant Cell Physiol, 2012, 53: 2127-2141.
doi: 10.1093/pcp/pcs154 |
| [32] |
Kaldenhoff R, Kölling A, Richter G. Regulation of the Arabidopsis thaliana aquaporin gene AthH2 (PIP1b). J Photochem Photobiol B: Biol, 1996, 36: 351-354.
doi: 10.1016/S1011-1344(96)07392-7 |
| [33] |
He W D, Gao J, Dou T X, Shao X H, Bi F C, Sheng O, Deng G M, Li C Y, Hu C H, Liu J H, Zhang S, Yang Q S, Yi G J. Early cold-induced peroxidases and aquaporins are associated with high cold tolerance in Dajiao (musa spp. ‘Dajiao’). Front Plant Sci, 2018, 9: 282.
doi: 10.3389/fpls.2018.00282 |
| [34] |
Ahamed A, Murai-Hatano M, Ishikawa-Sakurai J, Hayashi H, Kawamura Y, Uemura M. Cold stress-induced acclimation in rice is mediated by root-specific aquaporins. Plant Cell Physiol, 2012, 53: 1445-1456.
doi: 10.1093/pcp/pcs089 pmid: 22711693 |
| [35] |
Yamamori K, Ogasawara K, Ishiguro S, Koide Y, Takamure I, Fujino K, Sato Y, Kishima Y. Revision of the relationship between anther morphology and pollen sterility by cold stress at the booting stage in rice. Ann Bot, 2021, 128: 559-575.
doi: 10.1093/aob/mcab091 |
| [36] | Bots M, Feron R, Uehlein N, Weterings K, Kaldenhoff R, Mariani T. PIP1 and PIP2 aquaporins are differentially expressed during tobacco anther and stigma development. J Exp Bot, 2005, 56: 113-121. |
| [37] |
Tornroth-Horsefield S, Wang Y, Hedfalk K, Johanson U, Karlsson M, Tajkhorshid E, Neutze R, Kjellbom P. Structural mechanism of plant aquaporin gating. Nature, 2006, 439: 688-694.
doi: 10.1038/nature04316 |
| [38] | He X Y, Zhen H, Yin H Y, Chen F, Dong Y H, Zhang L F, Lu X Q, Zang J B, Ma W J, Mu P. High-throughput sequencing-based identification of miRNAs and their target mRNAs in wheat variety Qing Mai 6 under salt stress condition. Front Genet, 2021, 12: 724527. |
| [39] |
Duan F M, Ding J, Lee D S, Lu X L, Feng Y Q, Song W W. Overexpression of SoCYP85A1, a spinach cytochrome p450 gene in transgenic tobacco enhances root development and drought stress tolerance. Front Plant Sci, 2017, 8: 1909.
doi: 10.3389/fpls.2017.01909 |
| [40] |
Shen J, Diao W, Zhang L, Acharya B R, Zhang W. Secreted peptide PIP1 induces stomatal closure by activation of guard cell anion channels in Arabidopsis. Front Plant Sci, 2020, 11: 1029.
doi: 10.3389/fpls.2020.01029 |
| [41] |
Hendrick J P, Hartl F U. Molecular chaperone functions of heat-shock proteins. Annu Rev Biochem, 1993, 62: 349-384.
pmid: 8102520 |
| [42] | Banerjee A, Roychoudhury A. The role of aquaporins during plant abiotic stress responses. Plant Life Under Changing Environment. United Kingdom: Elsevier Academic Press, 2020. pp 643-661. |
| [43] | Kumar K, Mosa K A, Meselhy A G, Dhankher O P. Molecular insights into the plasma membrane intrinsic proteins roles for abiotic stress and metalloids tolerance and transport in plants. Indian J Plant Physiol, 2018, 23: 721-730. |
| [44] | de Paula Santos Martins C, Pedrosa A M, Du D, Gonçalves L P, Yu Q, Gmitter F G Jr, Costa M G C. Genome-wide characterization and expression analysis of major intrinsic proteins during abiotic and biotic stresses in sweet orange (Citrus sinensis L. Osb.). PLoS One, 2015, 10: e0138786. |
| [45] | Shafqat W, Jaskani M J, Maqbool R, Chattha W S, Ali Z, Naqvi S A, Haider M S, Khan I A, Vincent C I. Heat shock protein and aquaporin expression enhance water conserving behavior of citrus under water deficits and high temperature conditions. Environ Exp Bot, 2021, 181: 104270. |
| [46] |
Ma W, Xiao Y, Li Y, Hu P, Wang Z, Yang G, Wang J. Overexpression of CfPIP1-1, CfPIP1-2, and CfPIP1-4 genes of Catalpa fargesii in transgenic Arabidopsis thaliana under drought stress. J For Res, 2020, 32: 285-296.
doi: 10.1007/s11676-019-01082-w |
| [1] | 张一铎, 李国强, 孔忠新, 王玉泉, 李小利, 茹振钢, 贾海燕, 马正强. 基因聚合选育抗赤霉病小麦新品系百农4299[J]. 作物学报, 2022, 48(9): 2221-2227. |
| [2] | 冯子恒, 李晓, 段剑钊, 高飞, 贺利, 杨天聪, 戎亚思, 宋莉, 尹飞, 冯伟. 基于特征波段选择和机器学习的小麦白粉病高光谱遥感监测[J]. 作物学报, 2022, 48(9): 2300-2314. |
| [3] | 曹际玲, 曾青, 朱建国. 不同品种小麦灌浆期旗叶光合特性及光合基因表达对臭氧浓度升高的响应[J]. 作物学报, 2022, 48(9): 2339-2350. |
| [4] | 李永波, 崔德周, 黄琛, 隋新霞, 樊庆琦, 楚秀生. 高度特异性小麦ATG8抗体的研制及其在细胞自噬检测中的应用[J]. 作物学报, 2022, 48(9): 2390-2399. |
| [5] | 王云奇, 高福莉, 李傲, 郭同济, 戚留冉, 曾寰宇, 赵建云, 王笑鸽, 高国英, 杨佳鹏, 白金泽, 马亚欢, 梁月馨, 张睿. 小麦花后穗部温度变化规律及其与产量的关系[J]. 作物学报, 2022, 48(9): 2400-2408. |
| [6] | 杜启迪, 郭会君, 熊宏春, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 宋希云, 刘录祥. 小麦顶端小穗退化突变体asd1基因定位[J]. 作物学报, 2022, 48(8): 1905-1913. |
| [7] | 王沙沙, 黄超, 汪庆昌, 晁岳恩, 陈锋, 孙建国, 宋晓. 小麦籽粒大小相关基因TaGS2克隆及功能分析[J]. 作物学报, 2022, 48(8): 1926-1937. |
| [8] | 冯亚娟, 李廷轩, 蒲勇, 张锡洲. 不同镉积累类型小麦各器官镉积累分布规律及机理分析[J]. 作物学报, 2022, 48(7): 1761-1770. |
| [9] | 刘阿康, 马瑞琦, 王德梅, 王艳杰, 杨玉双, 赵广才, 常旭虹. 覆膜和补施氮肥对晚播冬小麦冬前植株生长及群体质量的影响[J]. 作物学报, 2022, 48(7): 1771-1786. |
| [10] | 王娟, 刘翼, 姚丹妤, 邹景伟, 肖世和, 孙果忠. 小麦生殖发育阶段对低温的敏感性鉴定[J]. 作物学报, 2022, 48(7): 1721-1729. |
| [11] | 张少华, 段剑钊, 贺利, 井宇航, 郭天财, 王永华, 冯伟. 基于无人机平台多模态数据融合的小麦产量估算研究[J]. 作物学报, 2022, 48(7): 1746-1760. |
| [12] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [13] | 郭星宇, 刘朋召, 王瑞, 王小利, 李军. 旱地冬小麦产量、氮肥利用率及土壤氮素平衡对降水年型与施氮量的响应[J]. 作物学报, 2022, 48(5): 1262-1272. |
| [14] | 付美玉, 熊宏春, 周春云, 郭会君, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 徐延浩, 刘录祥. 小麦矮秆突变体je0098的遗传分析与其矮秆基因定位[J]. 作物学报, 2022, 48(3): 580-589. |
| [15] | 冯健超, 许倍铭, 江薛丽, 胡海洲, 马英, 王晨阳, 王永华, 马冬云. 小麦籽粒不同层次酚类物质与抗氧化活性差异及氮肥调控效应[J]. 作物学报, 2022, 48(3): 704-715. |
|
||