欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (10): 2654-2664.doi: 10.3724/SP.J.1006.2024.41008
• 研究简报 • 上一篇
赵阳1,2( ), 李龙2, 杨进文1, 景蕊莲2, 孙黛珍1,*(), 王景一2,*()
), 李龙2, 杨进文1, 景蕊莲2, 孙黛珍1,*(), 王景一2,*()
ZHAO Yang1,2(), LI Long2, YANG Jin-Wen1, JING Rui-Lian2, SUN Dai-Zhen1,*(), WANG Jing-Yi2,*()
摘要:
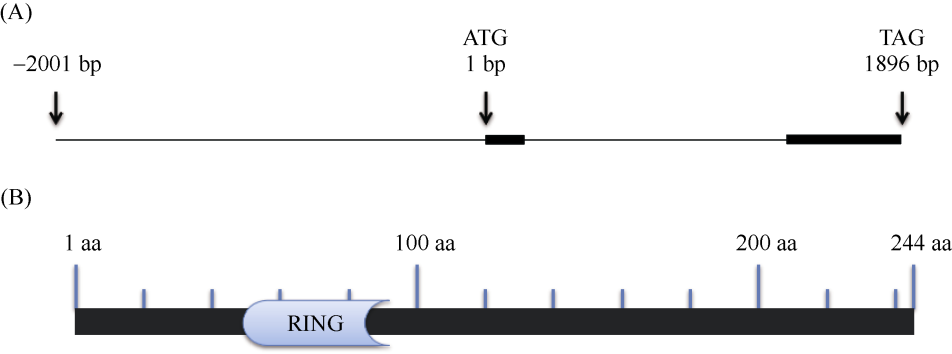
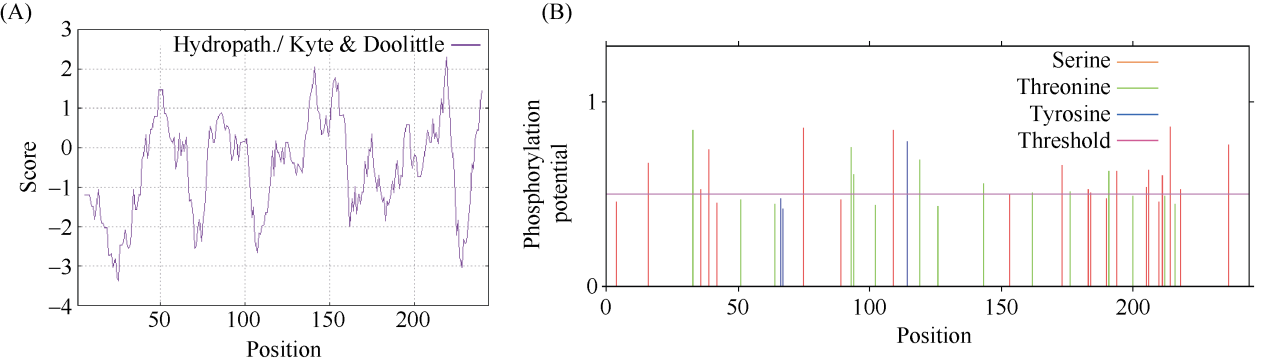
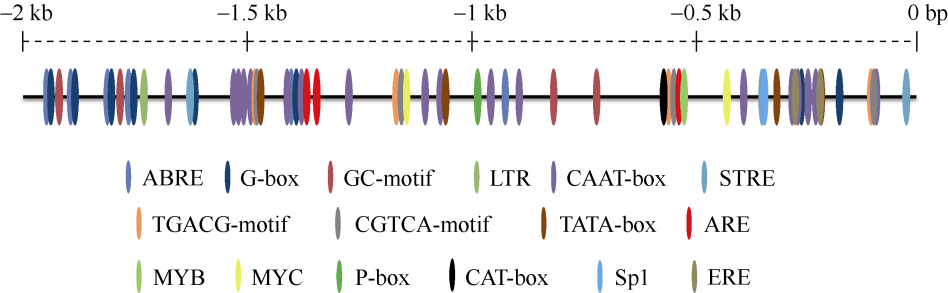
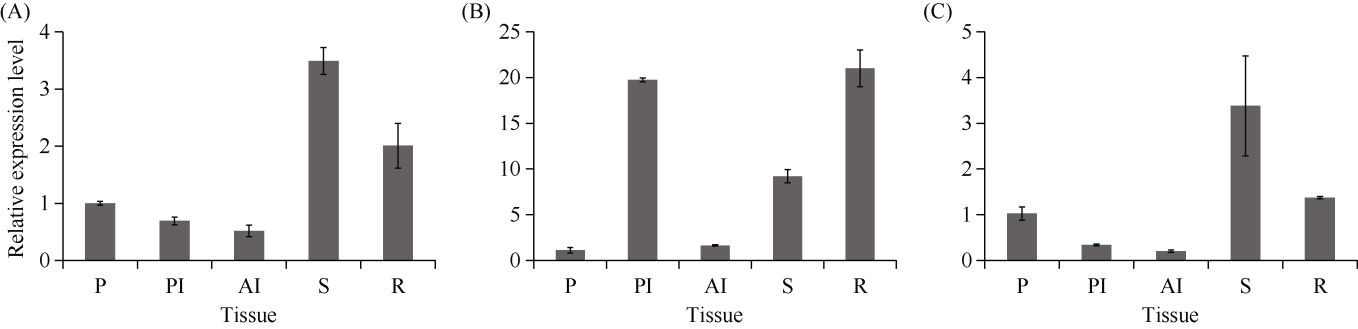
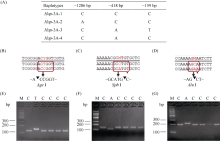
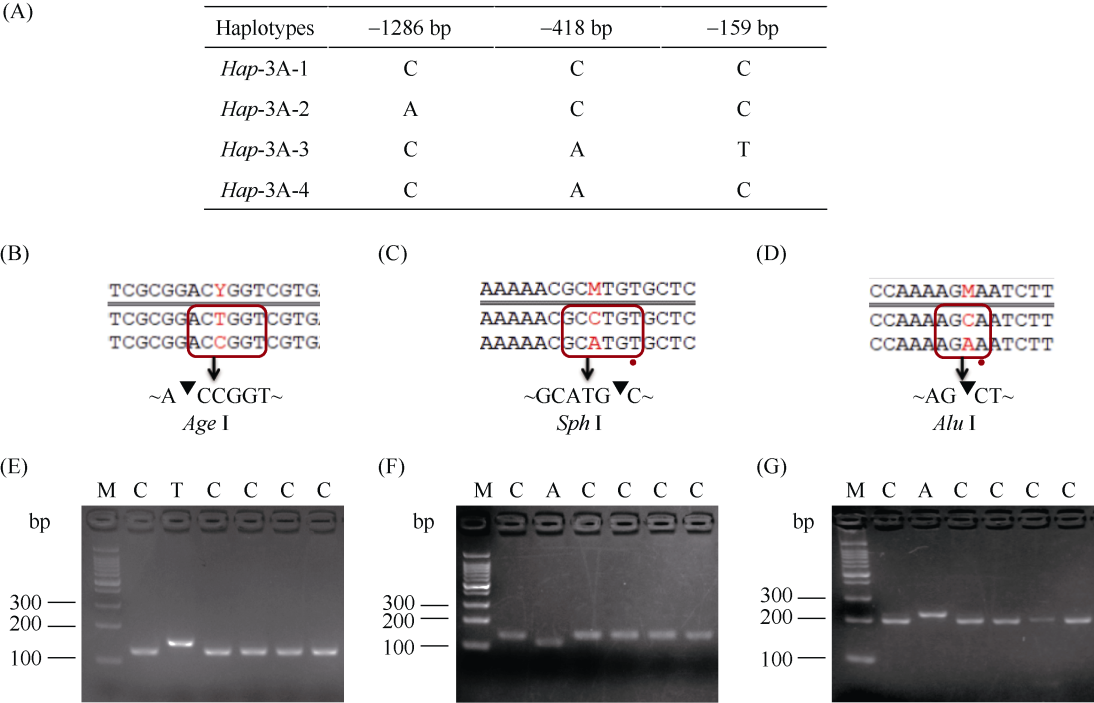
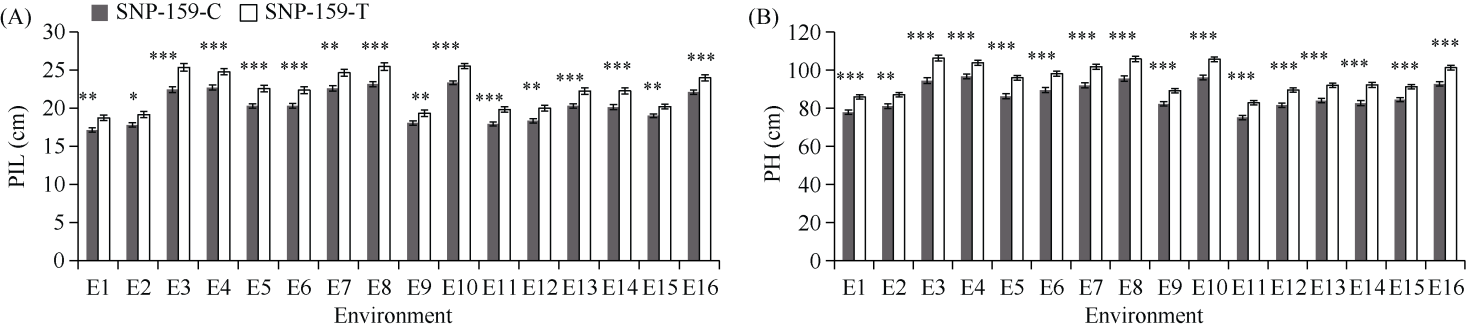
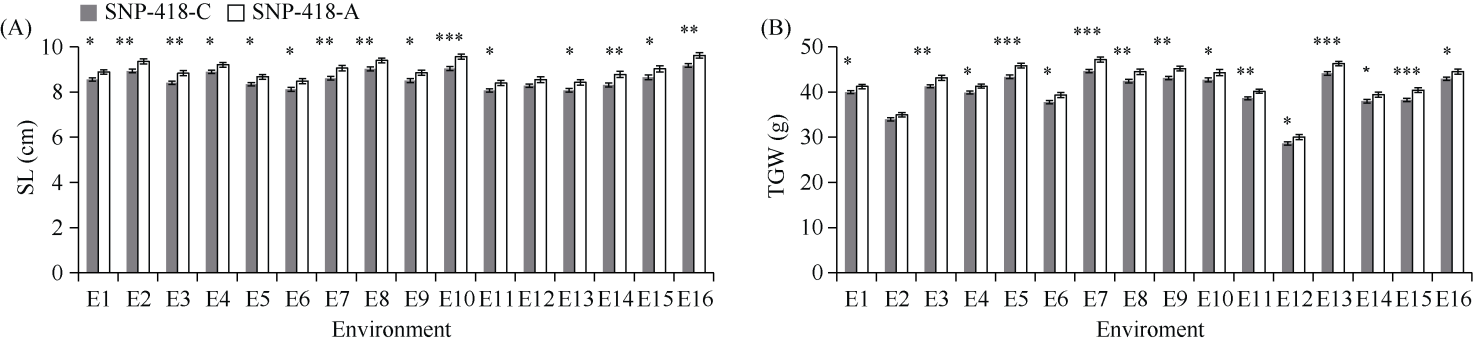
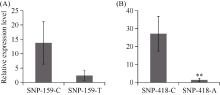
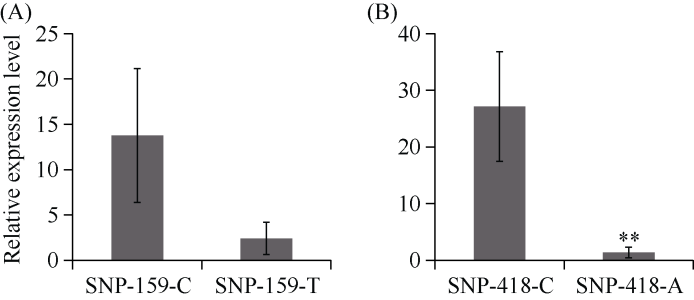
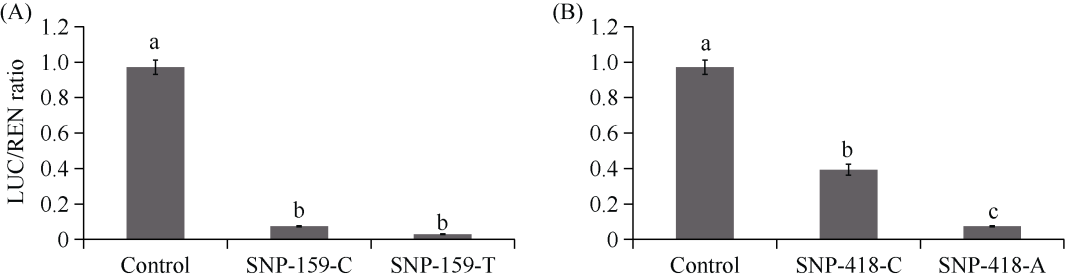
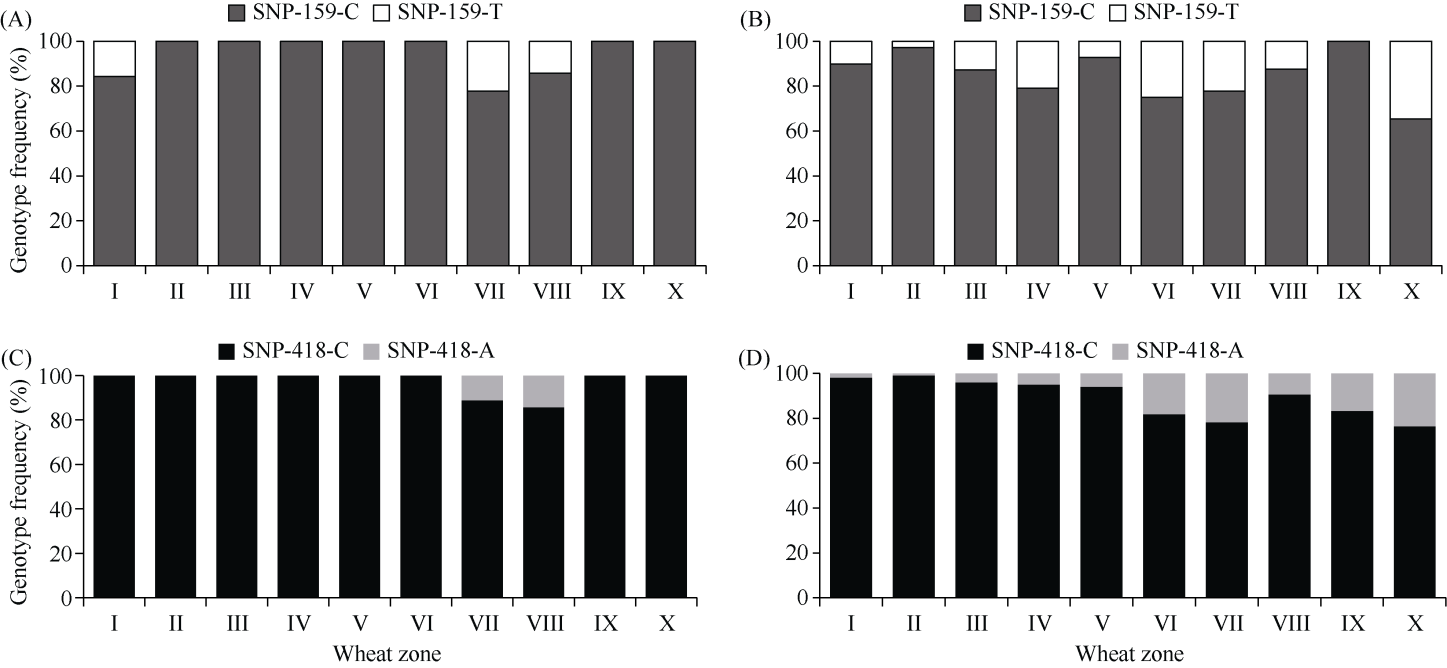
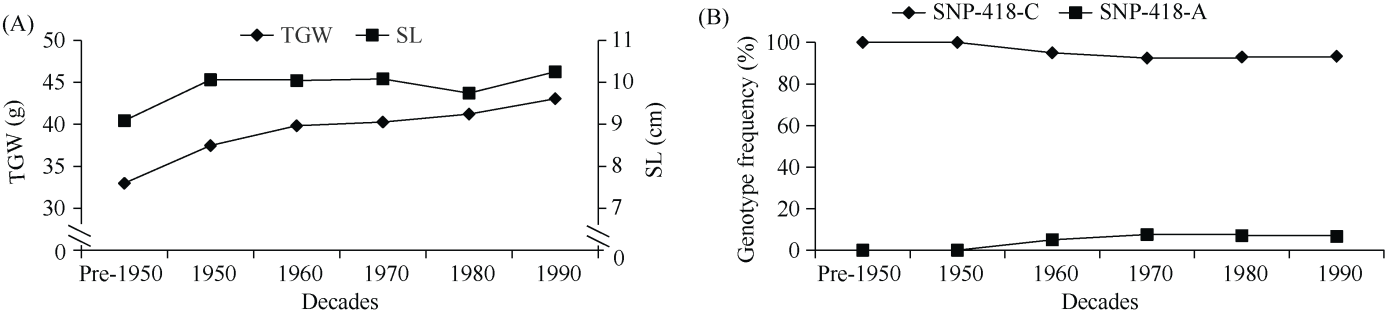
E3泛素连接酶SINA (seven in absentia)家族参与植物的生长发育、免疫共生并响应逆境胁迫。本研究克隆了位于小麦染色体3A上的TaSINA-3A基因, 其基因组序列长度为3897 bp, 其中启动子区为2001 bp, 基因序列为1896 bp, 编码244个氨基酸, 在50~91位氨基酸处有一个RING (really interesting new gene)保守结构域。RT-qPCR结果显示, TaSINA-3A基因在小麦的各个发育阶段的不同组织中均有表达。序列多态性分析表明, 在TaSINA-3A的启动子区存在33个SNP (single nucleotide polymorphism)位点, 编码区有4个SNP位点。基于启动子区的变异位点SNP-159、SNP-418和SNP-1286开发分子标记, 扫描小麦自然群体, 并与表型性状进行关联分析。结果表明,标记CAPS-159与多种环境下的株高和倒二节长显著相关, SNP-159-C是倒二节较短的矮秆等位变异, 标记dCAPS-418分别与多种环境下的穗长和千粒重显著关联, SNP-418-A是长穗、高千粒重的优异等位变异; TaSINA-3A负调控穗和籽粒的发育, TaSINA-3A的表达可能受到MYC转录因子的抑制; 在我国小麦育种进程中SNP-418-A受到正向选择, 在育成品种中的频率逐步增加, 但尚未充分利用。研究结果为深入探讨小麦株高、倒二节长及产量的形成机制提供参考, 并为培育高产稳产广适新品种提供了基因资源。
| [1] | Erenstein O, Jaleta M, Mottaleb K A, Sonder K, Donovan J, Braun H J. Global Trends in Wheat Production, Consumption and Trade. Berlin: Springer International Publishing Cham, 2022. pp 47-66. |
| [2] |
Hedden P. The genes of the green revolution. Trends Genet, 2003, 19: 5-9.
doi: 10.1016/s0168-9525(02)00009-4 pmid: 12493241 |
| [3] |
Koppolu R, Schnurbusch T. Developmental pathways for shaping spike inflorescence architecture in barley and wheat. J Integr Plant Biol, 2019, 61: 278-295.
doi: 10.1111/jipb.12771 |
| [4] |
Cao S H, Xu D A, Hanif M, Xia X C, He Z H. Genetic architecture underpinning yield component traits in wheat. Theor Appl Genet, 2020, 133: 1811-1823.
doi: 10.1007/s00122-020-03562-8 pmid: 32062676 |
| [5] | Wang J P, Li C N, Mao X G, Wang J Y, Li L, Li J L, Fan Z P, Zhu Z, He L H, Jing R L. The wheat basic helix-loop-helix gene TabHLH123 positively modulates the formation of crown roots and is associated with plant height and 1000-grain weight under various conditions. J Exp Bot, 2023, 74: 2542-2555. |
| [6] |
姚琦馥, 周界光, 王健, 陈黄鑫, 杨瑶瑶, 刘倩, 闫磊, 王瑛, 周景忠, 崔凤娟, 蒋云, 马建. 小麦穗长QTL鉴定及其遗传分析. 中国农业科学, 2023, 56: 4814-4825.
doi: 10.3864/j.issn.0578-1752.2023.24.002 |
|
Yao Q F, Zhou J G, Wang J, Chen H X, Yang Y Y, Liu Q, Yan L, Wang Y, Zhou J Z, Cui F J, Jiang Y, Ma J. Identification and genetic analysis of QTL for spike length in wheat. Sci Agric Sin, 2023, 56: 4814-4825 (in Chinese with English abstract).
doi: 10.3864/j.issn.0578-1752.2023.24.002 |
|
| [7] |
董继梓, 陈林渠, 郭浩儒, 张梦宇, 刘志霄, 韩磊, 田赵飒爽, 徐宁浩, 郭庆杰, 黄振洁, 杨傲宇, 赵春华, 吴永振, 孙晗, 秦冉, 崔法. 小麦穗长主效QTL-qSl-2D的遗传和育种选择效应解析. 中国农业科学, 2023, 56: 3917-3930.
doi: 10.3864/j.issn.0578-1752.2023.20.001 |
| Dong J Z, Chen L Q, Guo H R, Zhang M Y, Liu Z X, Han L, Tian Z S S, Xu N H, Guo Q J, Huang Z J, Yang A Y, Zhao C H, Wu Y Z, Sun H, Qin R, Cui F. Analysis of genetic and breeding selection effects of a major QTL-qSl-2D for wheat spike length. Sci Agric Sin, 2023, 56: 3917-3930 (in Chinese with English abstract). | |
| [8] | 张有富. TaTAP46基因对小麦千粒重和籽粒大小的影响. 甘肃农业大学博士学位论文, 甘肃兰州, 2021. |
| Zhang Y F. Effect of TaTAP46 Gene on Thousand Kernel Weight and Kernel Size in Wheat. PhD Dissertation of Gansu Agricultural University, Lanzhou, Gansu, China, 2021 (in Chinese with English abstract). | |
| [9] | Vierstra R D. The ubiquitin-26S proteasome system at the nexus of plant biology. Nat Rev Mol Cell Biol, 2009, 10: 385-397. |
| [10] | Xu F Q, Xue H W. The ubiquitin-proteasome system in plant responses to environments. Plant Cell Environ, 2019, 42: 2931-2944. |
| [11] | 田爱梅, 于晖, 曹家树. 植物E3泛素连接酶的分类与功能. 中国细胞生物学学报, 2020, 42: 907-915. |
| Tian A M, Yu H, Cao J S. Classification and function of E3 ubiquitin ligase in plants. Chin J Cell Biol, 2020, 42: 907-915 (in Chinese with English abstract). | |
| [12] |
Sun J Q, Huang S Y, Lu Q, Li S, Zhao S Z, Zheng X J, Zhou Q, Zhang W X, Li J, Wang L L, Zhang K, Zheng W Y, Feng X Z, Liu B H, Kong F J, Xiang F N. UV-B irradiation-activated E3 ligase GmILPA1 modulates gibberellin catabolism to increase plant height in soybean. Nat Commun, 2023, 14: 6262.
doi: 10.1038/s41467-023-41824-3 pmid: 37805547 |
| [13] | Zhang J L, Li C N, Li L, Xi Y J, Wang J Y, Mao X G, Jing R L. RING finger E3 ubiquitin ligase gene TaAIRP2-1B controls spike length in wheat. J Exp Bot, 2023, 74: 5014-5025. |
| [14] | Wang J Y, Wang R T, Mao X G, Zhang J L, Liu Y N, Xie Q, Yang X Y, Chang X P, Li C N, Zhang X Y, Jing R L. RING finger ubiquitin E3 ligase gene TaSDIR1-4A contributes to determination of grain size in common wheat. J Exp Bot, 2020, 71: 5377-5388. |
| [15] | Zhang C Y, Hao Z Y, Ning Y S, Wang G L. SINA E3 ubiquitin ligases: versatile moderators of plant growth and stress response. Mol Plant, 2019, 12: 610-612. |
| [16] | Li L, Peng Z, Mao X G, Wang J Y, Chang X P, Reynolds M, Jing R L. Genome-wide association study reveals genomic regions controlling root and shoot traits at late growth stages in wheat. Ann Bot, 2019, 124: 993-1006. |
| [17] | Hao C Y, Wang L F, Ge H M, Dong Y C, Zhang X Y. Genetic diversity and linkage disequilibrium in Chinese bread wheat (Triticum aestivum L.) revealed by SSR markers. PLoS One, 2011, 6: e17279. |
| [18] | Li L, Mao X G, Wang J Y, Chang X P, Reynolds M, Jing R L. Genetic dissection of drought and heat-responsive agronomic traits in wheat. Plant Cell Environ, 2019, 42: 2540-2553. |
| [19] | Xie Q, Guo H S, Dallman G, Fang S, Weissman A M, Chua N H. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature, 2002, 419: 167-170. |
| [20] |
Nolan T M, Brennan B, Yang M R, Chen J N, Zhang M C, Li Z H, Wang X L, Bassham D C, Walley J, Yin Y H. Selective autophagy of BES1 mediated by DSK2 balances plant growth and survival. Dev Cell, 2017, 41: 33-46.
doi: S1534-5807(17)30166-1 pmid: 28399398 |
| [21] | Yang M R, Li C X, Cai Z Y, Hu Y M, Nolan T, Yu F F, Yin Y H, Xie Q, Tang G L, Wang X L. SINAT E3 ligases control the light-mediated stability of the brassinosteroid-activated transcription factor BES1 in Arabidopsis. Dev Cell, 2017, 41: 47-58. |
| [22] |
Ning Y S, Jantasuriyarat C, Zhao Q Z, Zhang H W, Chen S B, Liu J L, Liu L J, Tang S Y, Park C H, Wang X J, Liu X L, Dai L Y, Xie Q, Wang G L. The SINA E3 ligase OsDIS1 negatively regulates drought response in rice. Plant Physiol, 2011, 157: 242-255.
doi: 10.1104/pp.111.180893 pmid: 21719639 |
| [23] |
Bao Y, Wang C T, Jiang C M, Pan J, Zhang G B, Liu H, Zhang H X. The tumor necrosis factor receptor-associated factor (TRAF)-like family protein SEVEN IN ABSENTIA 2 (SINA2) promotes drought tolerance in an ABA-dependent manner in Arabidopsis. New Phytol, 2014, 202: 174-187.
doi: 10.1111/nph.12644 pmid: 24350984 |
| [24] | Xue Y H, Wang J Y, Mao X G, Li C N, Li L, Yang X, Hao C Y, Chang X P, Li R Z, Jing R L. Association analysis revealed that TaPYL4 genes are linked to plant growth related traits in multiple environments. Front Plant Sci, 2021, 12: 641087. |
| [25] |
范子培, 李龙, 史雨刚, 孙黛珍, 李超男, 景蕊莲. 小麦TabHLH112-2B基因克隆及每穗小穗数相关功能标记开发. 作物学报, 2024, 50: 403-413.
doi: 10.3724/SP.J.1006.2024.31016 |
| Fan Z P, Li L, Shi Y G, Sun D Z, Li C N, Jing R L. Cloning of TabHLH112-2B gene and development of its functional marker associated with the number of spikelets per spike in wheat. Acta Agron Sin, 2024, 50: 403-413 (in Chinese with English abstract). | |
| [26] | Gao C H, Qi S H, Liu K G, Li D, Jin C Y, Li Z W, Huang G Q, Hai J B, Zhang M, Chen M X. MYC2, MYC3, and MYC4 function redundantly in seed storage protein accumulation in Arabidopsis. Plant Physiol Biochem, 2016, 108: 63-70. |
| [27] | Wang W J, Fan Y H, Niu X L, Miao M, Kud J, Zhou B J, Zeng L R, Liu Y S, Xiao F M. Functional analysis of the seven in absentia ubiquitin ligase family in tomato. Plant Cell Environ, 2018, 41: 689-703. |
| [28] |
Kim J H, Khan I U, Lee C W, Kim D Y, Jang C S, Lim S D, Park Y C, Kim J H, Seo Y W. Identification and analysis of a differentially expressed wheat RING-type E3 ligase in spike primordia development during post-vernalization. Plant Cell Rep, 2021, 40: 543-558.
doi: 10.1007/s00299-020-02651-8 pmid: 33423075 |
| [29] | Lyu Q, Li L Q, Meng Y, Sun H M, Chen L P, Wang B X, Li X J. Wheat E3 ubiquitin ligase TaGW2-6A degrades TaAGPS to affect seed size. Plant Sci, 2022, 320: 111274. |
| [30] | 李淑敏. 泛素E3连接酶TaGW2调控小麦抗逆性的分子机制研究. 西北农林科技大学博士学位论文, 陕西杨凌, 2023. |
| Li S M. The Molecular Mechanism of E3 Ubiquitin Ligase TaGW2 in Regulating Wheat Stress Resistance. PhD Dissertation of Northwest Agriculture and Forestry University, Yangling, Shaanxi, China, 2023 (in Chinese with English abstract). |
| [1] | 叶靓, 朱叶琳, 裴琳婧, 张思颖, 左雪倩, 李正真, 刘芳, 谭静. 联合全基因组关联和转录组分析筛选玉米拟轮枝镰孢穗腐病的抗性候选基因[J]. 作物学报, 2024, 50(9): 2279-2296. |
| [2] | 禹海龙, 吴文雪, 裴星旭, 刘晓宇, 邓跟望, 李西臣, 甄士聪, 望俊森, 赵永涛, 许海霞, 程西永, 詹克慧. 小麦茎秆性状的转录组测序及全基因组关联分析[J]. 作物学报, 2024, 50(9): 2187-2206. |
| [3] | 艾莎, 李莎, 方治伟, 李论, 李甜甜, 高利芬, 陈利红, 肖华锋, 万人静, 闫多子, 武星廷, 彭海, 韩瑞玺, 周俊飞. 棉花MNP标记位点开发及其在DNA指纹图谱构建中的应用[J]. 作物学报, 2024, 50(9): 2267-2278. |
| [4] | 彭小爱, 卢茂昂, 张玲, 刘童, 曹磊, 宋有洪, 郑文寅, 何贤芳, 朱玉磊. 基于55K SNP芯片的小麦籽粒主要品质性状的全基因组关联分析[J]. 作物学报, 2024, 50(8): 1948-1960. |
| [5] | 裴法敬, 张文轩, 张晓, 王昕钰, 彭少兵, 米甲明. 长粒香型的超短生育期水稻新品系创制[J]. 作物学报, 2024, 50(7): 1684-1698. |
| [6] | 马艳明, 娄鸿耀, 王威, 孙娜, 颜国荣, 张胜军, 刘杰, 倪中福, 徐麟. 新疆冬小麦籽粒品质性状遗传差异与关联分析[J]. 作物学报, 2024, 50(6): 1394-1405. |
| [7] | 郑雪晴, 王兴荣, 张彦军, 龚佃明, 邱法展. 玉米果穗相关性状QTL定位及重要候选基因分析[J]. 作物学报, 2024, 50(6): 1435-1450. |
| [8] | 张红梅, 张威, 王琼, 贾倩茹, 孟珊, 熊雅文, 刘晓庆, 陈新, 陈华涛. 大豆籽粒Ve含量的全基因组关联分析[J]. 作物学报, 2024, 50(5): 1223-1235. |
| [9] | 曹松, 姚敏, 任睿, 贾元, 向星汝, 李文, 何昕, 刘忠松, 官春云, 钱论文, 熊兴华. 转录组结合区域关联分析挖掘油菜含油量积累的候选基因[J]. 作物学报, 2024, 50(5): 1136-1146. |
| [10] | 张力岚, 杨军, 王让剑. 茶树橙花叔醇和芳樟醇樱草糖苷含量全基因组关联分析及候选基因预测[J]. 作物学报, 2024, 50(4): 871-886. |
| [11] | 郝倩琳, 杨廷志, 吕新茹, 秦慧敏, 王亚林, 贾晨飞, 夏先春, 马武军, 徐登安. 小麦胚芽鞘长度QTL定位和GWAS分析[J]. 作物学报, 2024, 50(3): 590-602. |
| [12] | 王琼, 朱宇翔, 周密密, 张威, 张红梅, 陈新, 陈华涛, 崔晓艳. 大豆叶型性状全基因组关联分析与候选基因鉴定[J]. 作物学报, 2024, 50(3): 623-632. |
| [13] | 刘薇, 王玉斌, 李伟, 张礼凤, 徐冉, 王彩洁, 张彦威. 过量表达大豆异丙基苹果酸脱氢酶基因GmIPMDH促进植株开花和生长[J]. 作物学报, 2024, 50(3): 613-622. |
| [14] | 范子培, 李龙, 史雨刚, 孙黛珍, 李超男, 景蕊莲. 小麦TabHLH112-2B基因克隆及每穗小穗数相关功能标记开发[J]. 作物学报, 2024, 50(2): 403-413. |
| [15] | 刁现民, 王立伟, 智慧, 张俊, 李顺国, 程汝宏. 谷子中矮秆资源创制、遗传解析和育种利用[J]. 作物学报, 2024, 50(2): 265-279. |
|