欢迎访问作物学报,今天是
作物学报 ›› 2025, Vol. 51 ›› Issue (11): 2944-2957.doi: 10.3724/SP.J.1006.2025.54062
李万1,*( ), 常紫锐1, 卢瑶2, 沈日敏2, 赵永平1,*(), 白小东2,*()
), 常紫锐1, 卢瑶2, 沈日敏2, 赵永平1,*(), 白小东2,*()
LI Wan1,*(), CHANG Zi-Rui1, LU Yao2, SHEN Ri-Min2, ZHAO Yong-Ping1,*(), BAI Xiao-Dong2,*()
摘要:
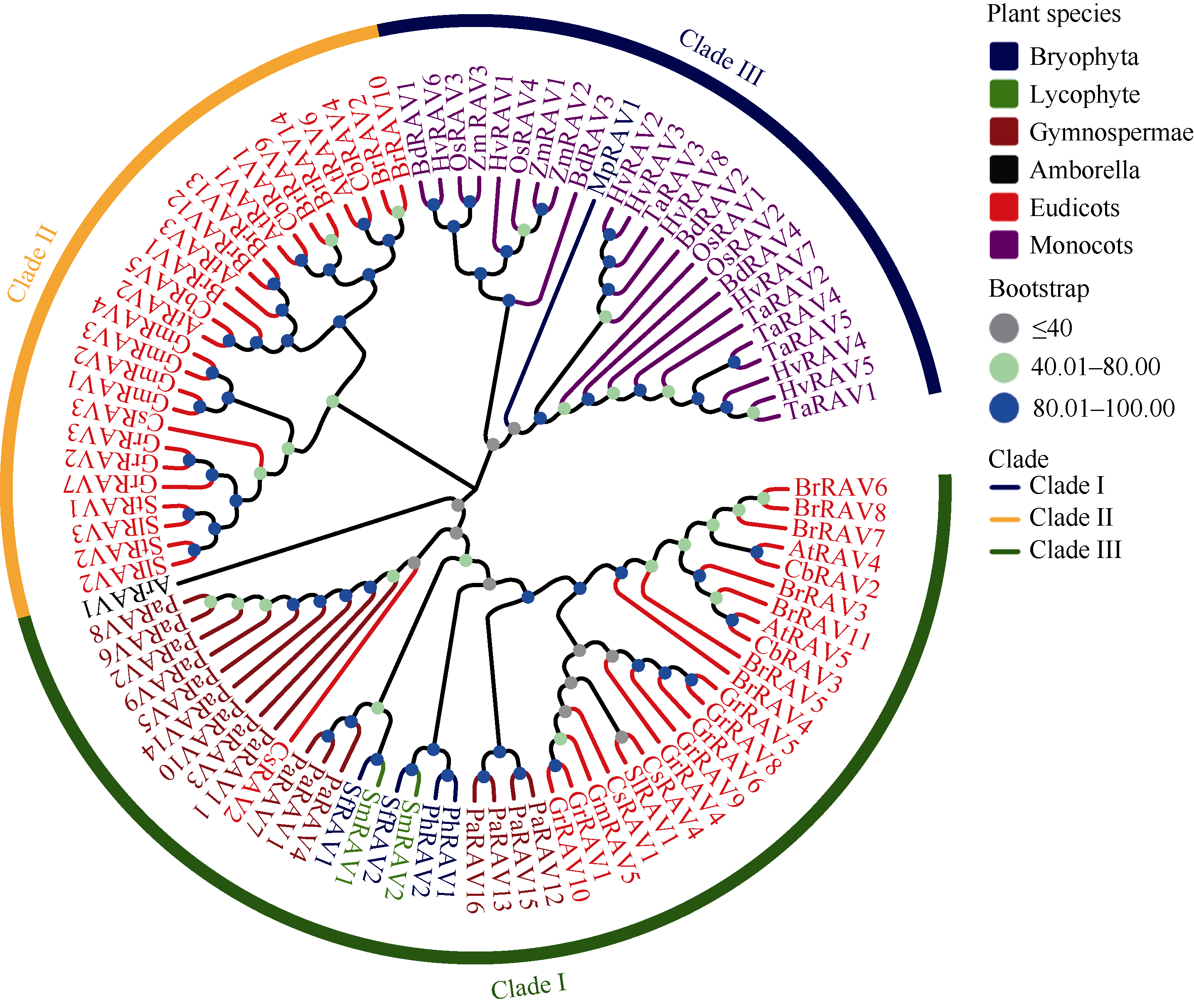
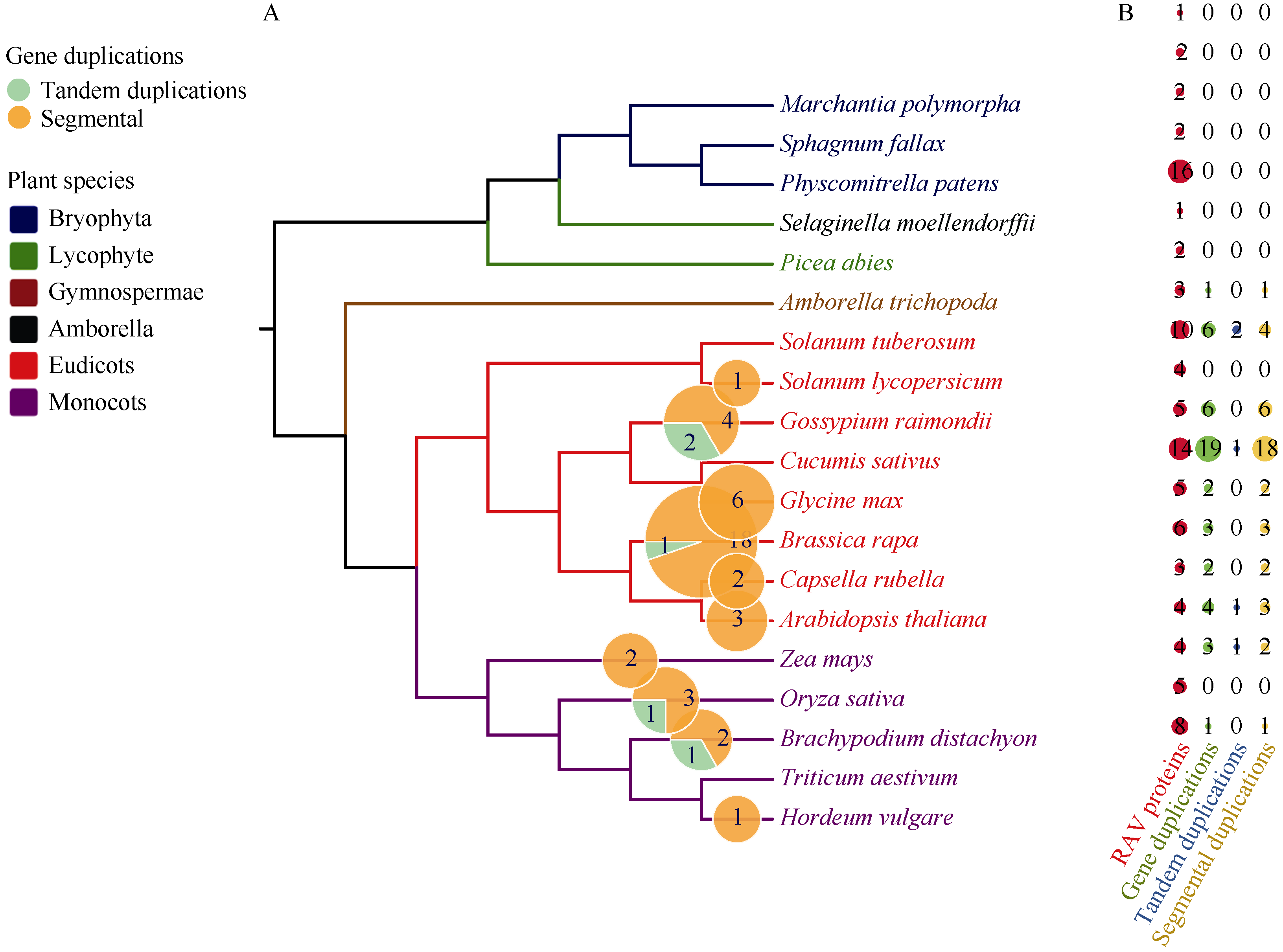
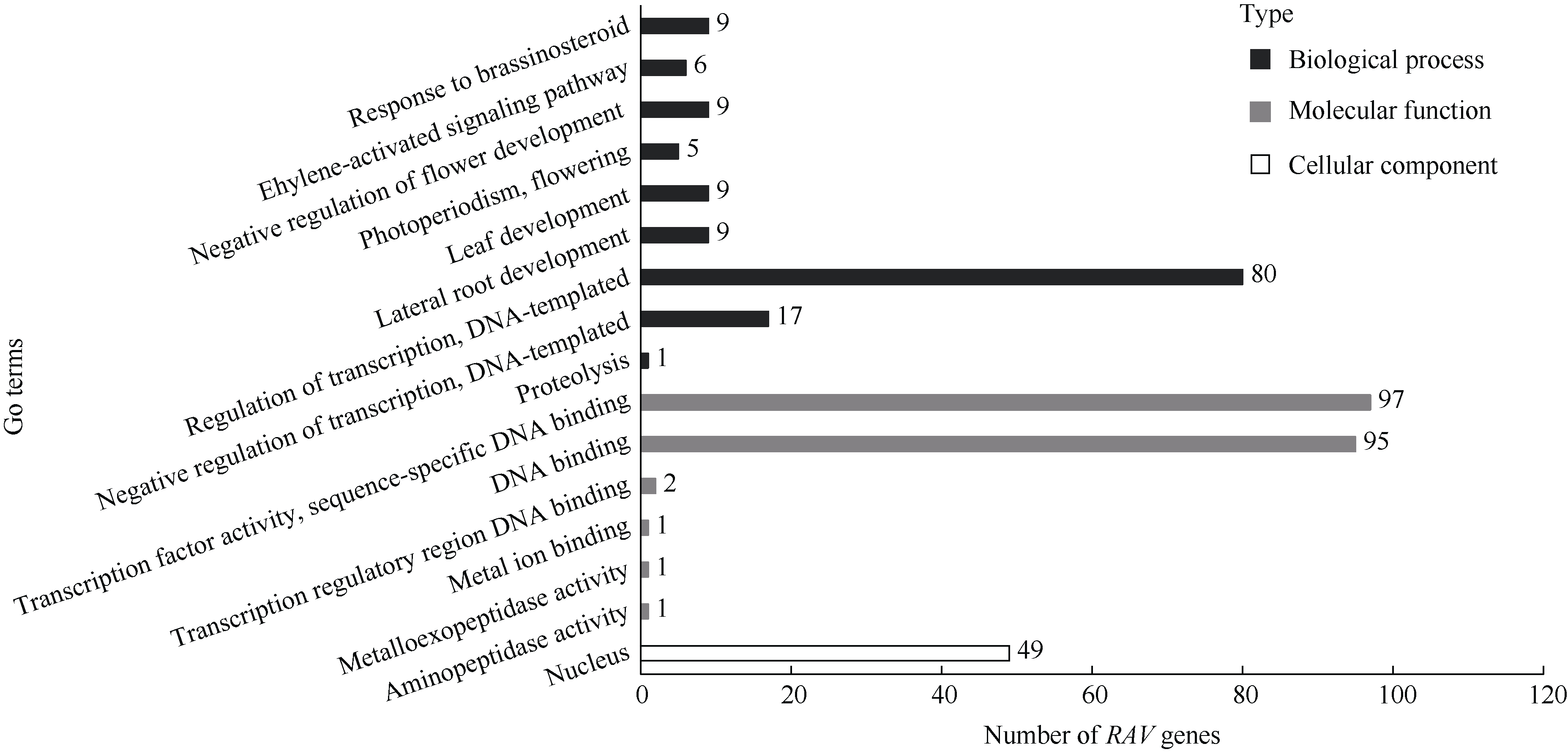
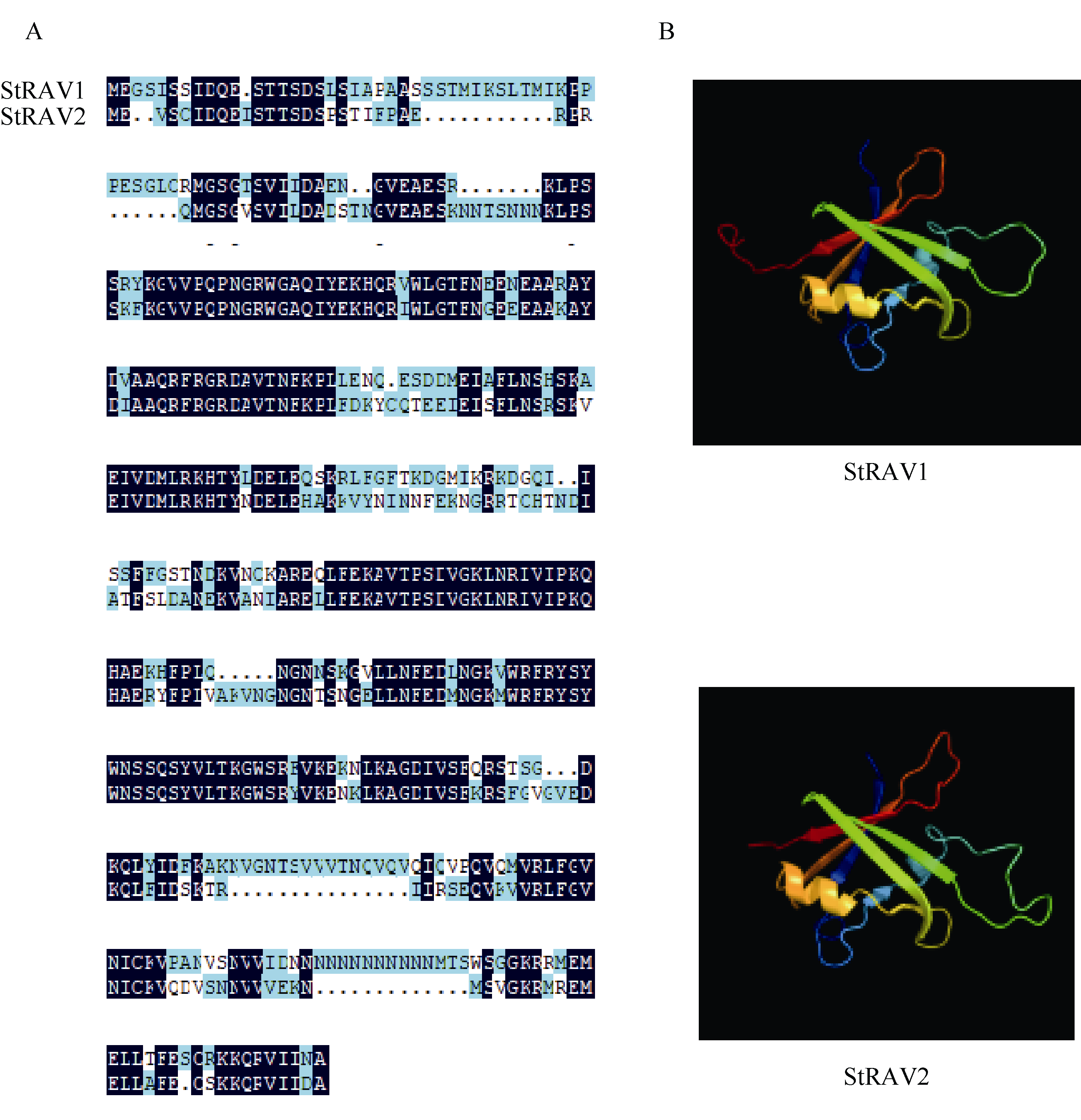
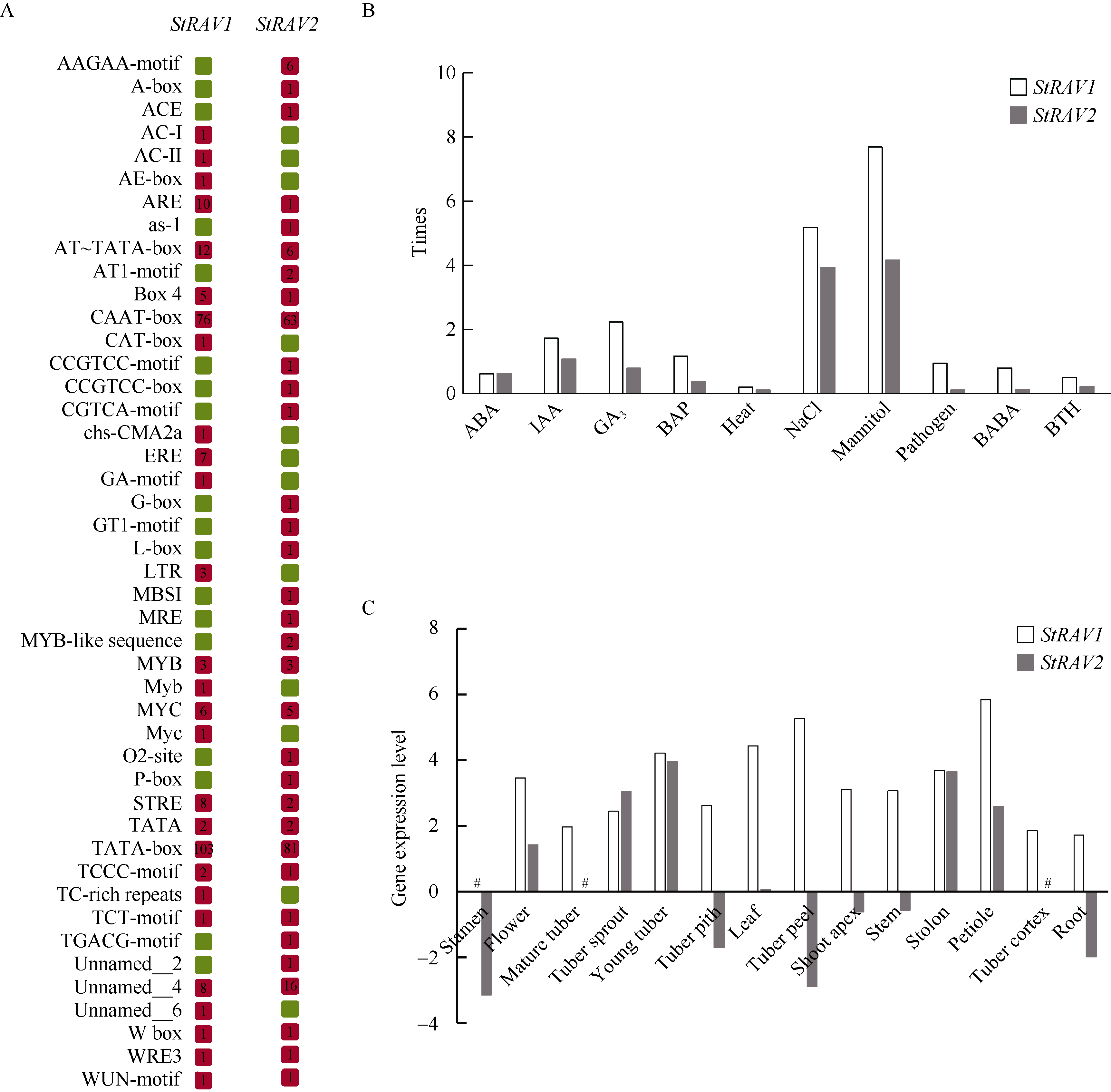
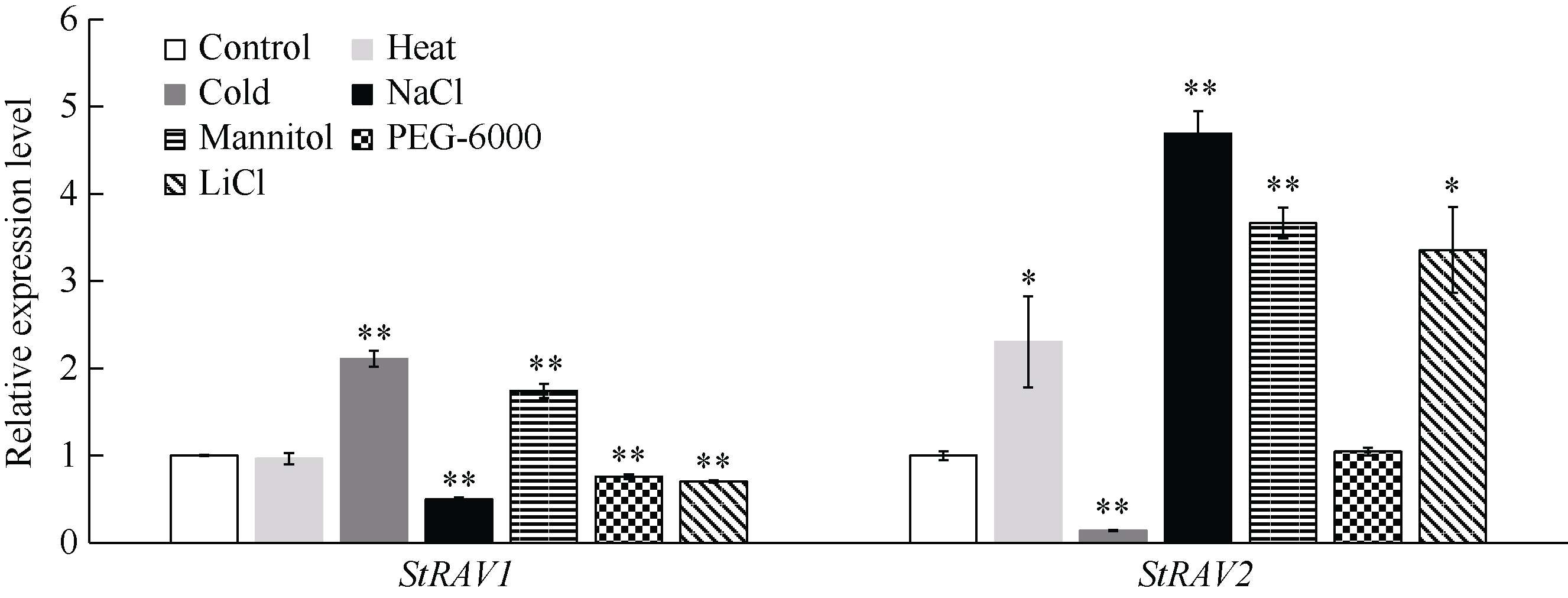
植物容易受到温度、高盐、干旱等各种环境因素的影响, 导致发育不良、降低产量。RAV家族是植物特有的, 与植物生长发育及生物和非生物胁迫抗性密切相关。本研究利用生物信息学方法和多个数据库, 对从藻类到被子植物的25种植物的RAV蛋白(RAVs)进行了鉴定和分析。结果表明, 裸子植物和被子植物中存在较多的RAVs, 苔藓植物和石松植物中存在少量的RAVs, 藻类植物中未发现RAVs。系统发育分析显示, RAVs被分为3个主要分支, Clade I包括从苔藓植物到被子植物的RAVs, 但单子叶植物除外。Clade II只含双子叶植物的RAVs。Clade III由单子叶植物和地钱的RAVs组成。此外, 通过对马铃薯RAVs的进一步分析表明, RAV家族成员数量虽然较少, 但在马铃薯响应非生物胁迫过程中具有重要作用, 也存在显著的功能分化。本研究结果详细鉴定和分析了25种植物中RAVs的性质和特征, 为进一步研究RAVs的功能提供了参考。
| [1] | Uçarlı C. Genome-wide identification and in silico gene expression analysis of the related to ABI3/VP1 (RAV) transcription factor family in barley (Hordeum vulgare L.). Biocell, 2021, 45: 1673-1685. |
| [2] | Zhou M L, Tang Y X, Wu Y M. Genome-wide analysis of AP2/ERF transcription factor family in Zea mays. Curr Bioinform, 2012, 7: 324-332. |
| [3] | Matías-Hernández L, Aguilar-Jaramillo A E, Marín-González E, Suárez-López P, Pelaz S. RAV genes: regulation of floral induction and beyond. Ann Bot, 2014, 114: 1459-1470. |
| [4] | Hu Y X, Wang Y X, Liu X F, Li J Y. Arabidopsis RAV1 is down-regulated by brassinosteroid and may act as a negative regulator during plant development. Cell Res, 2004, 14: 8-15. |
| [5] | Fu M J, Kang H K, Son S H, Kim S K, Nam K H. A subset of Arabidopsis RAV transcription factors modulates drought and salt stress responses independent of ABA. Plant Cell Physiol, 2014, 55: 1892-1904. |
| [6] |
Woo H R, Kim J H, Kim J, Kim J, Lee U, Song I J, Kim J H, Lee H Y, Nam H G, Lim P O. The RAV1 transcription factor positively regulates leaf senescence in Arabidopsis. J Exp Bot, 2010, 61: 3947-3957.
doi: 10.1093/jxb/erq206 pmid: 20826506 |
| [7] |
Lee S C, Choi D S, Hwang I S, Hwang B K. The pepper oxidoreductase CaOXR1 interacts with the transcription factor CaRAV1 and is required for salt and osmotic stress tolerance. Plant Mol Biol, 2010, 73: 409-424.
doi: 10.1007/s11103-010-9629-0 pmid: 20333442 |
| [8] |
Zhao L, Luo Q L, Yang C L, Han Y P, Li W B. A RAV-like transcription factor controls photosynthesis and senescence in soybean. Planta, 2008, 227: 1389-1399.
doi: 10.1007/s00425-008-0711-7 pmid: 18297307 |
| [9] | Lu Q Y, Zhao L, Li D M, Hao D Q, Zhan Y, Li W B. A GmRAV ortholog is involved in photoperiod and sucrose control of flowering time in soybean. PLoS One, 2014, 9: e89145. |
| [10] | Li C W, Su R C, Cheng C P, Sanjaya, You S J, Hsieh T H, Chao T C, Chan M T. Tomato RAV transcription factor is a pivotal modulator involved in the AP2/EREBP-mediated defense pathway. Plant Physiol, 2011, 156: 213-227. |
| [11] |
Li W, Chen Y, Ye M H, Lu H B, Wang D D, Chen Q. Evolutionary history of the C-repeat binding factor/dehydration-responsive element-binding 1 (CBF/DREB1) protein family in 43 plant species and characterization of CBF/DREB1 proteins in Solanum tuberosum. BMC Evol Biol, 2020, 20: 142.
doi: 10.1186/s12862-020-01710-8 pmid: 33143637 |
| [12] |
Singh V K, Jain M, Garg R. Genome-wide analysis and expression profiling suggest diverse roles of GH3 genes during development and abiotic stress responses in legumes. Front Plant Sci, 2014, 5: 789.
doi: 10.3389/fpls.2014.00789 pmid: 25642236 |
| [13] | Guo B J, Wei Y F, Xu R B, Lin S, Luan H Y, Lyu C, Zhang X Z, Song X Y, Xu R G. Genome-wide analysis of APETALA2/ ethylene-responsive factor (AP2/ERF) gene family in barley (Hordeum vulgare L.). PLoS One, 2016, 11: e0161322. |
| [14] |
Charfeddine M, Saïdi M N, Charfeddine S, Hammami A, Gargouri Bouzid R. Genome-wide analysis and expression profiling of the ERF transcription factor family in potato (Solanum tuberosum L.). Mol Biotechnol, 2015, 57: 348-358.
doi: 10.1007/s12033-014-9828-z pmid: 25491236 |
| [15] | Zhao S P, Xu Z S, Zheng W J, Zhao W, Wang Y X, Yu T F, Chen M, Zhou Y B, Min D H, Ma Y Z, et al. Genome-wide analysis of the RAV family in soybean and functional identification of GmRAV-03 involvement in salt and drought stresses and exogenous ABA treatment. Front Plant Sci, 2017, 8: 905. |
| [16] | Jin J P, Zhang H, Kong L, Gao G, Luo J C. PlantTFDB 3.0: a portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Res, 2014, 42: D1182-D1187. |
| [17] |
Bhattacharyya S, Pattanaik S, Maiti I B. Intron-mediated enhancement of gene expression in transgenic plants using chimeric constructs composed of the Peanut chlorotic streak virus (PClSV) promoter-leader and the antisense orientation of PClSV orf vii (p7R). Planta, 2003, 218: 115-124.
doi: 10.1007/s00425-003-1078-4 pmid: 12883884 |
| [18] |
Zhang C, Kong N N, Cao M X, Wang D D, Chen Y, Chen Q. Evolutionary significance of amino acid permease transporters in 17 plants from Chlorophyta to Angiospermae. BMC Genomics, 2020, 21: 391.
doi: 10.1186/s12864-020-6729-3 pmid: 32503414 |
| [19] |
Dossa K, Wei X, Li D H, Fonceka D, Zhang Y X, Wang L H, Yu J Y, Liao B S, Diouf D, Cissé N, et al. Insight into the AP2/ERF transcription factor superfamily in sesame and expression profiling of DREB subfamily under drought stress. BMC Plant Biol, 2016, 16: 171.
doi: 10.1186/s12870-016-0859-4 pmid: 27475988 |
| [20] |
Rogozin I B, Carmel L, Csuros M, Koonin E V. Origin and evolution of spliceosomal introns. Biol Direct, 2012, 7: 11.
doi: 10.1186/1745-6150-7-11 pmid: 22507701 |
| [21] | Roy S W, Penny D. Patterns of intron loss and gain in plants: intron loss-dominated evolution and genome-wide comparison of O. sativa and A. thaliana. Mol Biol Evol, 2007, 24: 171-181. |
| [22] | Kang J M, Ju H L, Sohn W M, Na B K. Characterization of the biochemical properties of two methionine aminopeptidases of Cryptosporidium parvum. Parasitol Int, 2012, 61: 707-710. |
| [23] |
Rice D W, Alverson A J, Richardson A O, Young G J, Sanchez-Puerta M V, Munzinger J, Barry K, Boore J L, Zhang Y, DePamphilis C W, et al. Horizontal transfer of entire genomes via mitochondrial fusion in the angiosperm Amborella. Science, 2013, 342: 1468-1473.
doi: 10.1126/science.1246275 pmid: 24357311 |
| [24] | Mittal A, Jiang Y W, Ritchie G L, Burke J J, Rock C D. AtRAV1 and AtRAV2 overexpression in cotton increases fiber length differentially under drought stress and delays flowering. Plant Sci, 2015, 241: 78-95. |
| [25] |
Sohn K H, Lee S C, Jung H W, Hong J K, Hwang B K. Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt stress tolerance. Plant Mol Biol, 2006, 61: 897-915.
doi: 10.1007/s11103-006-0057-0 pmid: 16927203 |
| [26] |
Zhuang J, Sun C C, Zhou X R, Xiong A S, Zhang J. Isolation and characterization of an AP2/ERF-RAV transcription factor BnaRAV-1-HY15 in Brassica napus L. Huyou15. Mol Biol Rep, 2011, 38: 3921-3928.
doi: 10.1007/s11033-010-0508-1 pmid: 21116861 |
| [27] | Hurst L D. The Ka/Ks ratio: diagnosing the form of sequence evolution. Trends Genet, 2002, 18: 486. |
| [28] | Conesa A, Götz S. Blast2GO: a comprehensive suite for functional analysis in plant genomics. Int J Plant Genomics, 2008, 2008: 619832. |
| [29] | Wei Y X, Chang Y L, Zeng H Q, Liu G Y, He C Z, Shi H T. RAV transcription factors are essential for disease resistance against cassava bacterial blight via activation of melatonin biosynthesis genes. J Pineal Res, 2018, 64: e12454. |
| [30] |
支添添, 周舟, 陈纪鹏, 韩成云. 甘蓝型油菜酪氨酸代谢关键基因FAH的克隆、功能鉴定和表达分析. 生物技术通报, 2023, 39(10): 115-127.
doi: 10.13560/j.cnki.biotech.bull.1985.2023-0270 |
| Zhi T T, Zhou Z, Chen J P, Han C Y. Cloning, functional identification and expression analysis of FAH, a key gene for tyrosine metabolism in Brassica napus L. Biotechnol Bull, 2023, 39(10): 115-127 (in Chinese with English abstract). | |
| [31] |
Titarenko E, Rojo E, León J, Sánchez-Serrano J J. Jasmonic acid-dependent and-independent signaling pathways control wound-induced gene activation in Arabidopsis thaliana. Plant Physiol, 1997, 115: 817-826.
pmid: 9342878 |
| [32] | Li B B, Zhao Y J, Wang S, Zhang X H, Wang Y W, Shen Y, Yuan Z H. Genome-wide identification, gene cloning, subcellular location and expression analysis of SPL gene family in P. granatum L. BMC Plant Biol, 2021, 21: 400. |
| [33] | 解盛, 李国旗, 宋立肖, 谢博勋, 王雅芳, 刘星. 罗布麻CesA基因家族的生物信息学分析. 广西植物, 2021, 41: 522-534. |
| Xie S, Li G Q, Song L X, Xie B X, Wang Y F, Liu X. Bioinformatics analysis of Apocynum venetum CesA gene family. Guihaia, 2021, 41: 522-534 (in Chinese with English abstract). | |
| [34] | Zhou L J, Wang Y X, Wang Y G, Song A P, Jiang J F, Chen S M, Ding B Q, Guan Z Y, Chen F D. Transcription factor CmbHLH16 regulates petal anthocyanin homeostasis under different lights in Chrysanthemum. Plant Physiol, 2022, 190: 1134-1152. |
| [35] | Zhu Y S, Wang Y C, Jiang H Y, Liu W J, Zhang S H, Hou X K, Zhang S S, Wang N, Zhang R, Zhang Z Y, et al. Transcriptome analysis reveals that PbMYB61 and PbMYB308 are involved in the regulation of lignin biosynthesis in pear fruit stone cells. Plant J, 2023, 116: 217-233. |
| [36] | Feng C Z, Chen Y, Wang C, Kong Y H, Wu W H, Chen Y F. Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development. Plant J, 2014, 80: 654-668. |
| [37] | Lee H Y, Byeon Y, Back K. Melatonin as a signal molecule triggering defense responses against pathogen attack in Arabidopsis and tobacco. J Pineal Res, 2014, 57: 262-268. |
| [1] | 贾小霞, 齐恩芳, 文国宏, 马胜, 黄伟, 吕和平, 李建武, 曲亚英, 丁宁. 中早熟马铃薯‘陇薯20号’高效再生体系建立及抗草铵膦种质创制[J]. 作物学报, 2025, 51(9): 2285-2294. |
| [2] | 卓峰琦, 唐振三, 雷雨俊, 程李香, 赵甜甜, 吕汰, 杨晨, 张峰. 基于烹饪方式及回生温度筛选低升糖马铃薯品种(系)[J]. 作物学报, 2025, 51(9): 2538-2546. |
| [3] | 朱锦程, 杨秋华, 程李香, 李文丽, 石明明, 李惠霞, 张峰. 马铃薯抗南方根结线虫种质资源筛选及相关生理反应分析[J]. 作物学报, 2025, 51(9): 2307-2317. |
| [4] | 尹丽娜, 张锐, 陈国欢, 白磊, 李俊, 郭华春, 杨芳. 不同马铃薯品种块茎创伤愈合能力的比较[J]. 作物学报, 2025, 51(9): 2399-2411. |
| [5] | 马娟娥, 姚有华, 姚晓华, 吴昆仑, 崔永梅. 青稞HvERF039基因的克隆及功能研究[J]. 作物学报, 2025, 51(9): 2341-2357. |
| [6] | 邵顺伟, 陈卓, 兰振东, 蔡兴奎, 邹华芬, 李晨曦, 唐景华, 朱熙, 张彧, 董建科, 金辉, 宋波涛. 基于BSA-seq技术的块茎芽眼深度QTL定位分析[J]. 作物学报, 2025, 51(7): 1725-1735. |
| [7] | 闫知兰, 赵芹, 常甜达, 王一鸣, 王碧辉, 王鹏, 黄春国, 张会, 王利祥, 郝晓鹏, 赵波. 豆科作物AOX基因鉴定及其在普通菜豆响应非生物胁迫中的表达模式研究[J]. 作物学报, 2025, 51(7): 1769-1783. |
| [8] | 李秋云, 李世贵, 范军亮, 刘昊天, 赵晓斌, 吕硕, 王艳浩, 岳云, 张宁, 司怀军. 离子锌和纳米锌对马铃薯生理特性、产量及品质的影响[J]. 作物学报, 2025, 51(7): 1838-1849. |
| [9] | 杨海洋, 吴林宣, 李博纹, 石翰峰, 袁禧龙, 刘金朝, 蔡海荣, 陈诗怡, 郭涛, 王慧. 基于QTL定位发现的OsWRI3调控水稻种子的落粒性[J]. 作物学报, 2025, 51(7): 1712-1724. |
| [10] | 凤舞剑, 冼晓青, 张新钵, 曹丹, 强承魁. 基于转录组和AlphaFold对稻瘟菌经典效应蛋白和水稻受体的快速鉴定[J]. 作物学报, 2025, 51(6): 1480-1488. |
| [11] | 杨双, 白磊, 郭华春, 缪亚生, 李俊. 马铃薯叶片表皮毛形态特征、类型与发育过程[J]. 作物学报, 2025, 51(6): 1582-1598. |
| [12] | 徐杰, 夏露露, 唐振三, 李文丽, 赵甜甜, 程李香, 张峰. 马铃薯块茎蒸制和烘焙后嗅味品质分析[J]. 作物学报, 2025, 51(5): 1409-1420. |
| [13] | 王林, 陈晓雨, 张文梦龙, 汪思琦, 程冰云, 程靖秋, 潘锐, 张文英. 大麦HvMYB2分子特性及响应干旱胁迫的功能分析[J]. 作物学报, 2025, 51(4): 873-887. |
| [14] | 赵喜娟, 张帆, 刘圣宣, 覃骏, 陈惠兰, 林原, 罗红兵, 刘易, 宋波涛, 胡新喜, 王恩爽. 4种马铃薯内源激素提取方法优化及其在块茎解除休眠过程中的含量分析[J]. 作物学报, 2025, 51(4): 1050-1060. |
| [15] | 方应浩, 周波, 陈茹梅, 杨文竹, 秦慧民. 基于RNA-seq和PER-seq联合分析探究ZmHDZ6表达调控网络[J]. 作物学报, 2025, 51(4): 958-968. |
|
||