欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (3): 687-702.doi: 10.3724/SP.J.1006.2023.24042
齐燕妮1( ), 李闻娟1, 赵丽蓉2, 李雯2, 王利民1, 谢亚萍1, 赵玮1, 党照1, 张建平1,*()
), 李闻娟1, 赵丽蓉2, 李雯2, 王利民1, 谢亚萍1, 赵玮1, 党照1, 张建平1,*()
QI Yan-Ni1(), LI Wen-Juan1, ZHAO Li-Rong2, LI Wen2, WANG Li-Min1, XIE Ya-Ping1, ZHAO Wei1, DANG Zhao1, ZHANG Jian-Ping1,*()
摘要:
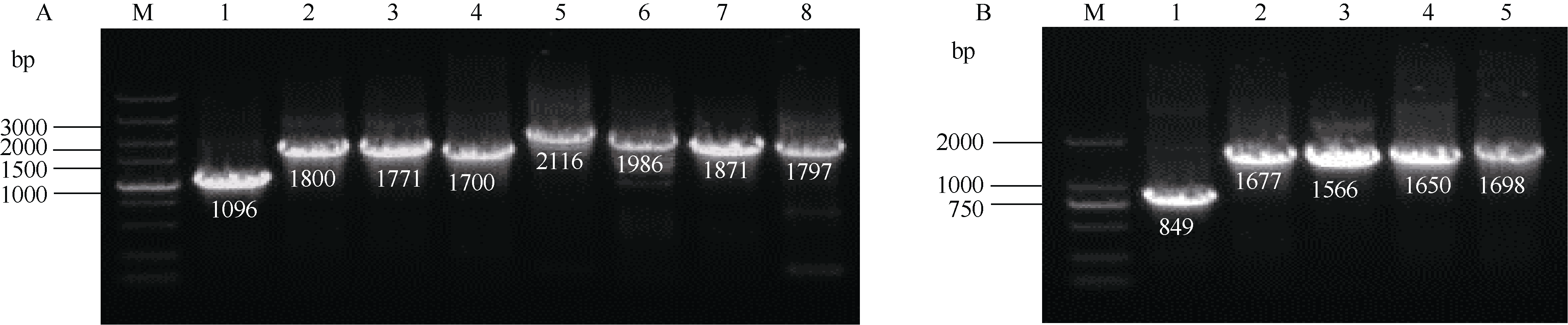
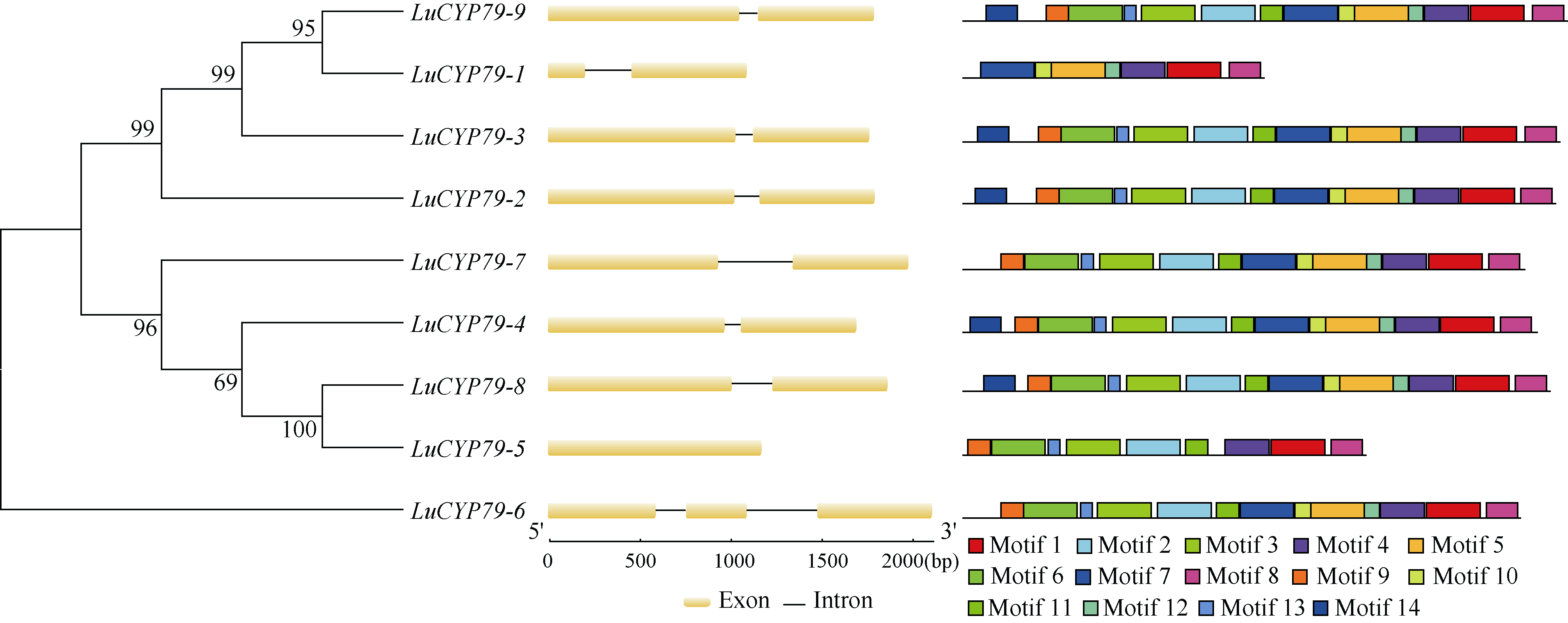
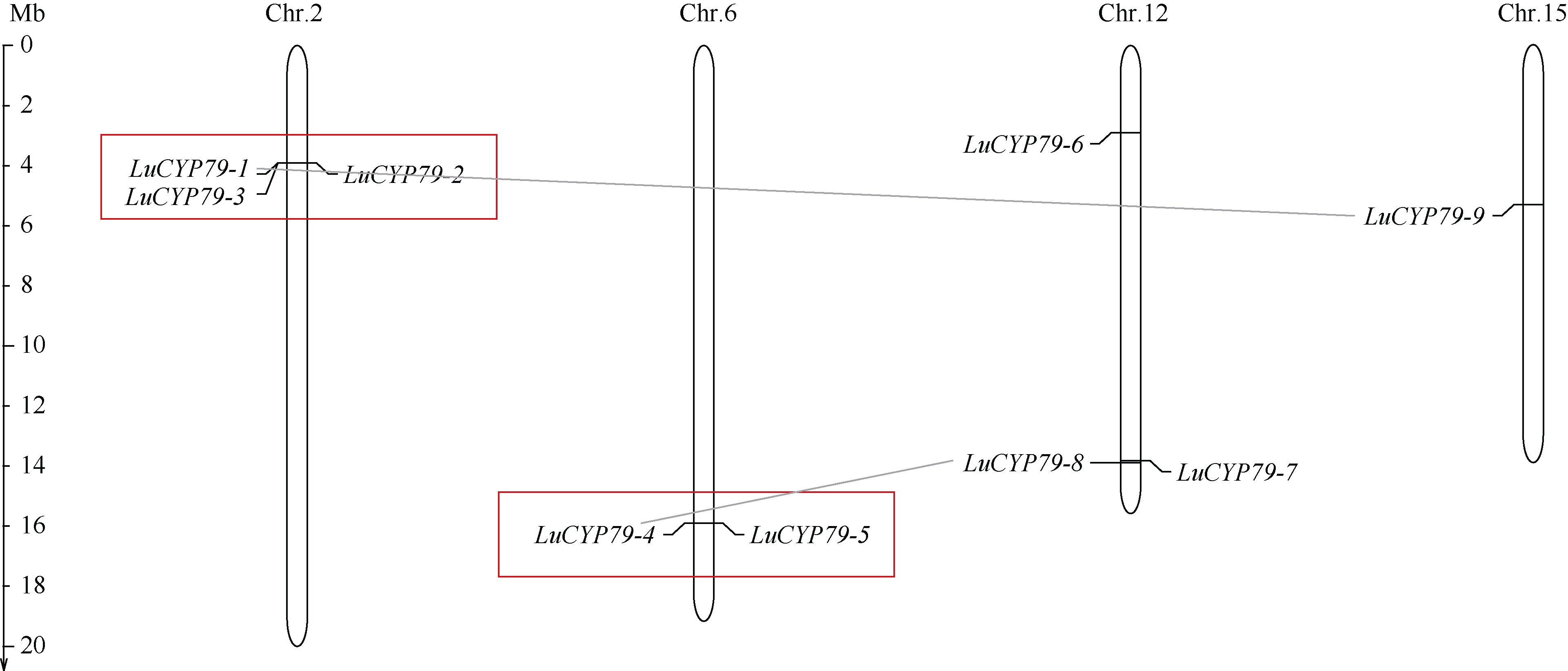
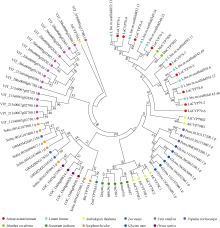
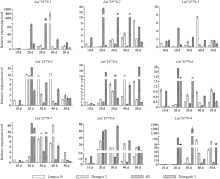
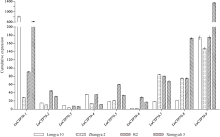
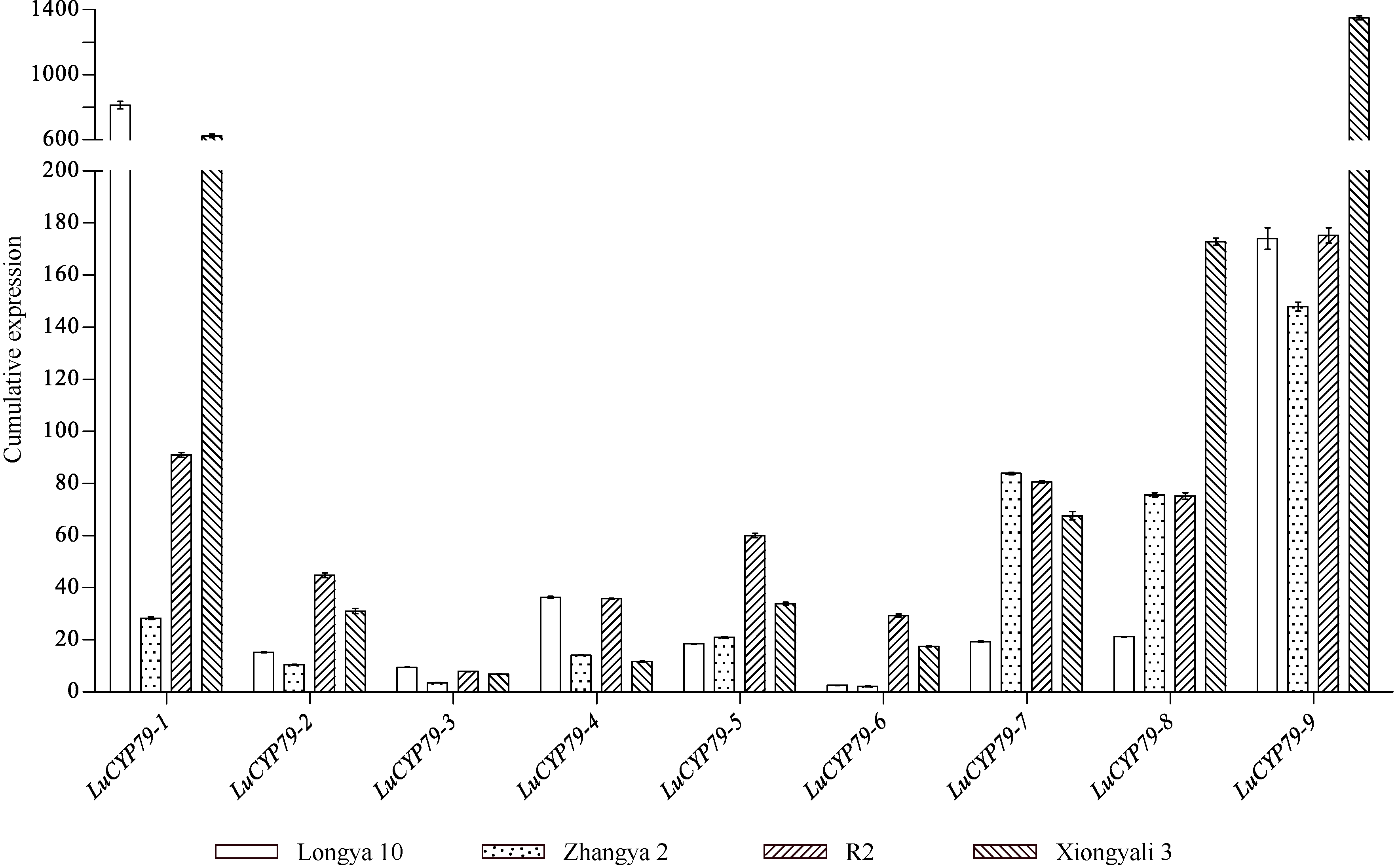
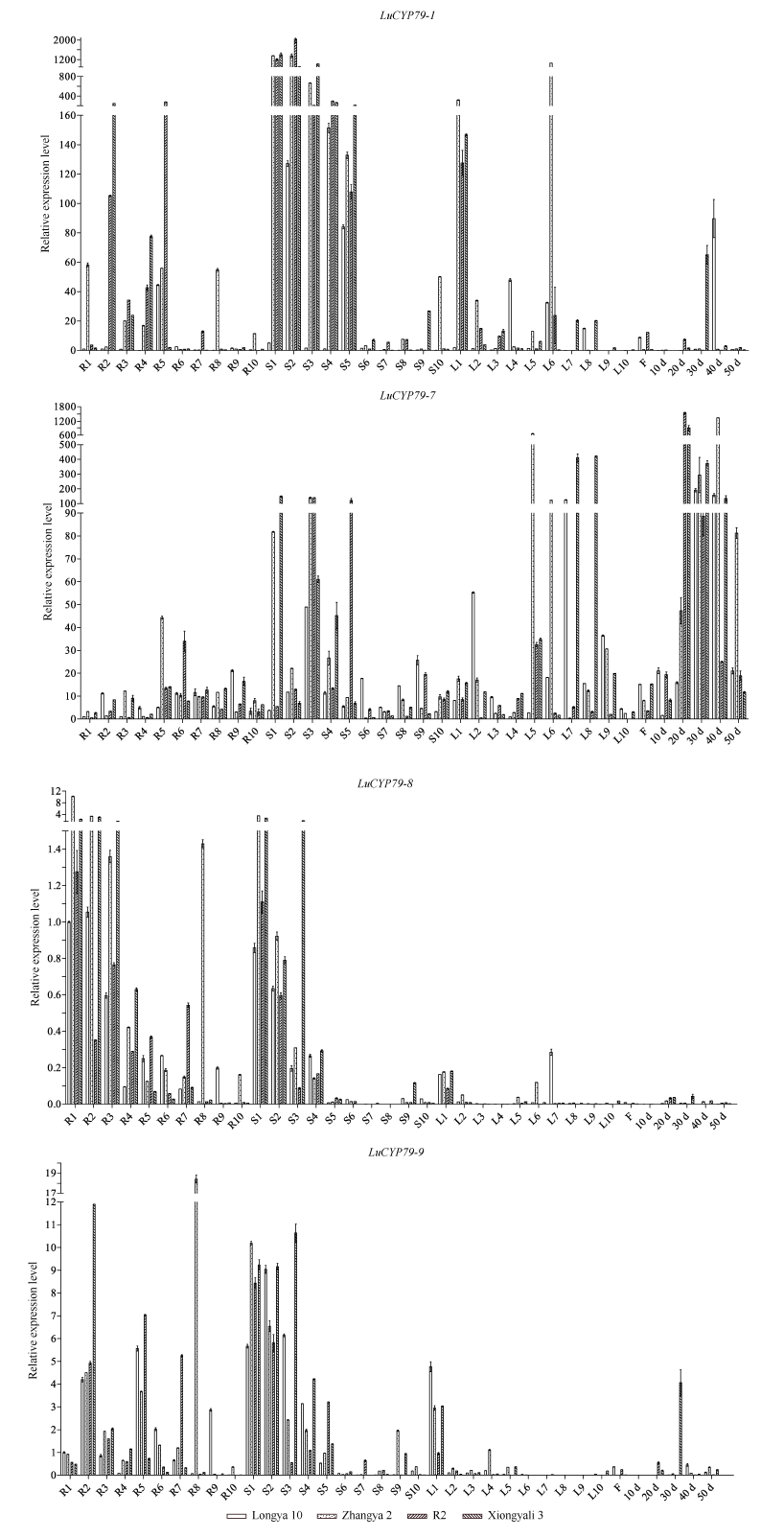
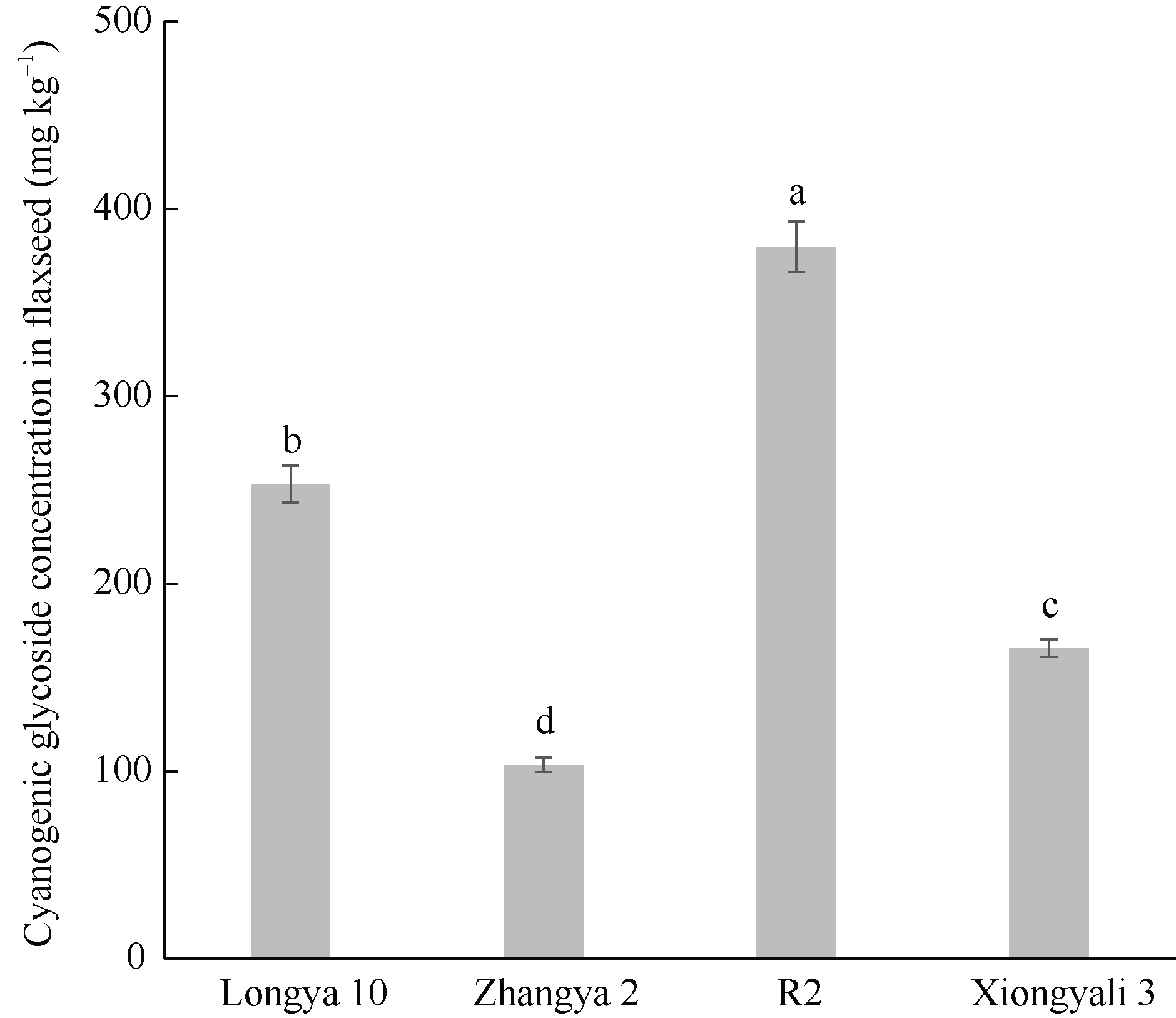
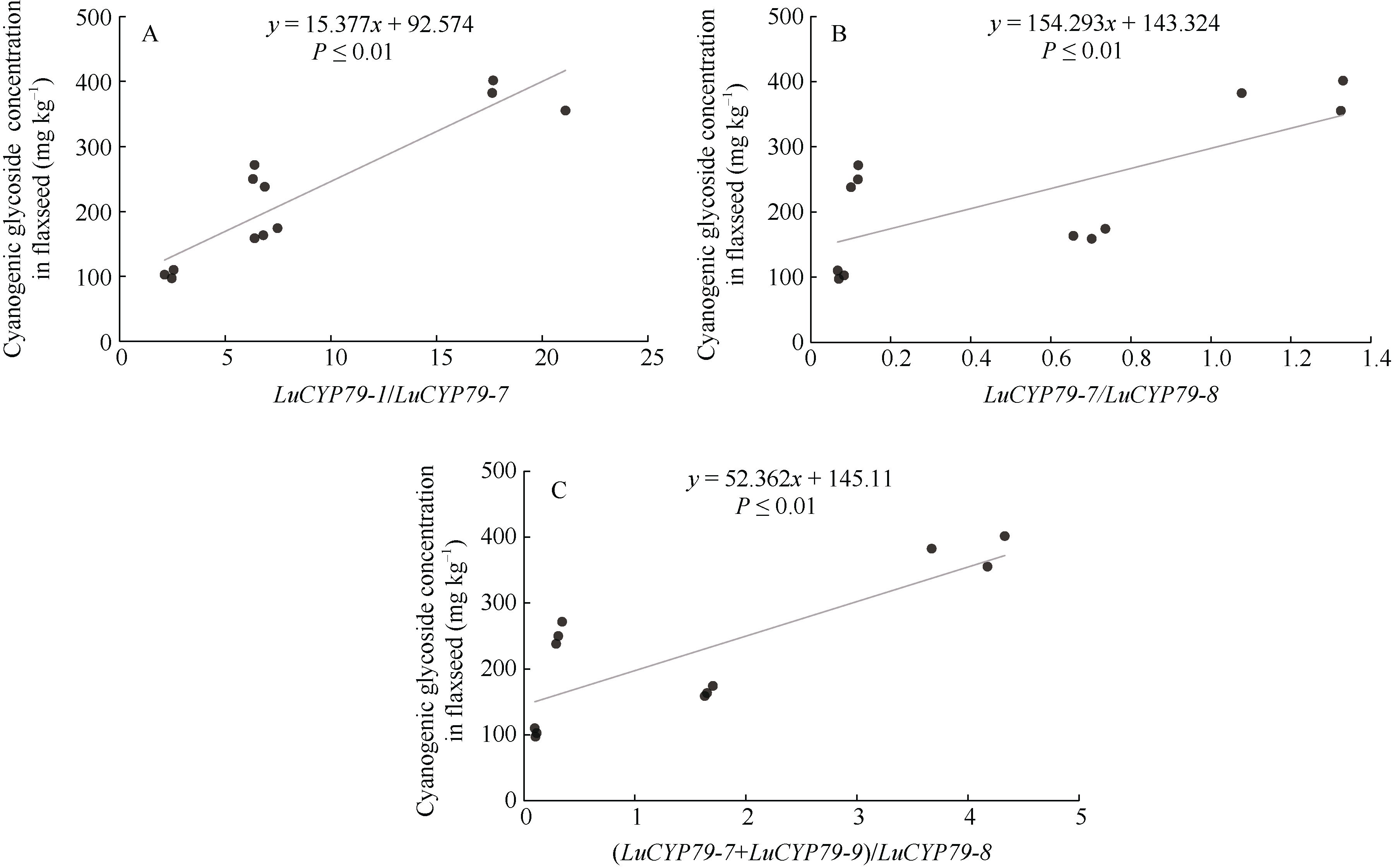
CYP79蛋白是生氰糖苷合成关键酶, 但关于亚麻CYP79基因的研究鲜有报道。本研究对包括亚麻在内的9种作物的CYP79基因家族进行了鉴定, 并分析了亚麻CYP79基因的序列特征、复制事件、共线性关系、系统进化、顺式作用元件及表达模式。结果表明, 在亚麻、白亚麻、毛果杨、木薯、芝麻、高粱、大豆、葡萄及水稻中分别鉴定到9、9、3、2、5、7、6、16和4个CYP79家族成员; 系统进化分析显示, CYP79基因的进化具有物种特异性; LuCYP79不均匀分布在4条染色体上, 具有1~3个外显子, 其启动子区含大量激素与逆境响应相关元件; 共克隆到8个亚麻CYP79基因的全长DNA序列及5个成员的全长cDNA序列; LuCYP79蛋白序列长度为282~565 aa, 等电点为5.84~9.14, 分子量为31.56~62.86 kD, 均为亲水性蛋白, 定位于内质网; 共有5对基因发生复制事件, 占全部基因的77.8%, 全部经历了强烈的纯化选择, 其中LuCYP79-1和LuCYP79-9在拟南芥和木薯中均具有同源基因。表达分析表明, LuCYP79家族成员具有组织特异性, 且各成员在不同遗传背景下表达模式不同, 其中LuCYP79-1、LuCYP79-7、LuCYP79-8和LuCYP79-9在4个品种中的表达差异显著。相关分析表明, 50 d时的LuCYP79-1/LuCYP79-7与成熟亚麻籽中生氰糖苷含量呈极显著正相关, 20 d时的LuCYP79-7/LuCYP79-8及(LuCYP79-7+LuCYP79-9)/LuCYP79-8分别与成熟亚麻籽中生氰糖苷含量呈极显著正相关, 初步推测其可能是亚麻籽生氰糖苷合成的关键基因。研究结果对进一步阐明亚麻CYP79蛋白的功能具有积极意义, 并为培育低生氰糖苷亚麻品种提供了理论参考。
| [1] |
Zohary D. Monophyletic vs. polyphyletic origin of the crops on which agriculture was founded in the near east. Genet Resour Crop Evol, 1999, 46: 133-142.
doi: 10.1023/A:1008692912820 |
| [2] |
Oomah B D. Flaxseed as a functional food source. J Sci Food Agric, 2001, 81: 889-894.
doi: 10.1002/jsfa.898 |
| [3] |
Turner T D, Mapiye C, Aalhus J L, Beaulieu A D, Patience J F, Zijlstra R T. Flaxseed fed pork: n-3 fatty acidenrichment and contribution to dietary recommendations. Meat Sci, 2014, 96: 541-547.
doi: 10.1016/j.meatsci.2013.08.021 pmid: 24012977 |
| [4] | Goyal A, Sharma V, Upadhyay N, Gill S, Sihag M. Flax and flaxseed oil: an ancient medicine & modernfunctional food. J Food Sci Technol, 2014, 9: 1633-1653. |
| [5] |
Oomah B D, Mazza G, Kenaschuk E O. Cyanogenic compounds in flaxseed. J Agric Food Chem, 1992, 40: 1346-1348.
doi: 10.1021/jf00020a010 |
| [6] | Conn E E. Cyanogenic glycosides. Biochem Plants, 1981, 7: 479-500. |
| [7] |
Nelson L. Acute cyanide toxicity mechanisms and manifestations. J Emerg Nurs, 2006, 32: 8-10.
pmid: 16860675 |
| [8] | Ganjewala D, Kumar S, Devi A, Ambika K. Advances in cyanogenic glycosides biosynthesis and analyses in plants. Acta Biol Szegediensis, 2010, 54: 1-14. |
| [9] |
Sibbesen O, Koch B, Halkier B A, Moller B L. Cytochrome P-450TYR is a multifunctional heme-thiolate enzyme catalyzing the conversion of L-tyrosine to p-hydroxyphenylacetaldehyde oxime in the biosynthesis of the cyanogenic glucoside dhurrin in Sorghum bicolor (L.) Moench. J Biol Chem, 1995, 270: 3506-3511.
doi: 10.1074/jbc.270.8.3506 pmid: 7876084 |
| [10] |
Wang C W, Dissing M M, Agerbirk N, Crocoll C, Halkier B A. Characterization of Arabidopsis CYP79C1 and CYP79C2 by glucosinolate pathway engineering in Nicotiana benthamiana shows substrate specificity toward a range of aliphatic and aromatic amino acids. Front Plant Sci, 2020, 11: 57.
doi: 10.3389/fpls.2020.00057 |
| [11] |
Andersen M D, Busk P K, Svendsen I, Møller B L. Cytochromes P-450 from cassava (Manihot esculenta Crantz) catalyzing the first steps in the biosynthesis of the cyanogenic glucosides linamarin and lotaustralin: cloning, functional expression in Pichia pastoris, and substrate specificity of the isolated recombinant enzymes. J Biol Chem, 2000, 275: 1966-1975.
doi: 10.1074/jbc.275.3.1966 pmid: 10636899 |
| [12] |
Irmisch S, McCormick A C, Boeckler G A, Schmidt A, Reichelt M, Schneider B, Block K, Schnitzler J, Gershenzon J, Unsicker S B, Köllner T G. Two herbivore-induced cytochrome P450 enzymes CYP79D6 and CYP79D7 catalyze the formation of volatile aldoximes involved in poplar defense. Plant Cell, 2013, 25: 4737-4754.
doi: 10.1105/tpc.113.118265 |
| [13] |
Irmisch S, Zeltner P, Handrick V, Gershenzon J, Köllner T G. The maize cytochrome P450 CYP79A61 produces phenylacetaldoxime and indole-3-acetaldoxime in heterologous systems and might contribute to plant defense and auxin formation. BMC Plant Biol, 2015, 15: 128-139.
doi: 10.1186/s12870-015-0526-1 pmid: 26017568 |
| [14] |
Bak S, Nielsen H L, Halkier B A. The presence of CYP79 homologues in glucosinolate-producing plants shows evolutionary conservation of the enzymes in the conversion of amino acid to aldoxime in the biosynthesis of cyanogenic glucosides and glucosinolates. Plant Mol Biol, 1998, 38: 725-734.
pmid: 9862490 |
| [15] |
Petersen B L, Andreasson E, Bak S, Agerbirk N, Halkier B A. Characterization of transgenic Arabidopsis thaliana with metabolically engineered high levels of p-hydroxybenzylglucosinolate. Planta, 2001, 212: 612-618.
pmid: 11525519 |
| [16] |
Reintanz B, Lehnen M, Reichelt M, Gershenzon J, Kowalczyk M, Sandberg G, Godde M, Uhl R, Palme K. Bus, a bushy Arabidopsis CYP79F1 knockout mutant with abolished synthesis of short-chain aliphatic glucosinolates. Plant Cell, 2001, 13: 351-367.
doi: 10.1105/tpc.13.2.351 pmid: 11226190 |
| [17] |
Chen S X, Glawischnig E, Jørgensen K, Naur P, Jørgensen B, Olsen C E, Hansen C H, Rasmussen H, Pickett J A, Halkier B A. CYP79F1 and CYP79F2 have distinct functions in the biosynthesis of aliphatic glueosinolates in Arabidopsis. Plant J, 2003, 33: 923-937.
doi: 10.1046/j.1365-313X.2003.01679.x |
| [18] |
Bak S, Paquette S M, Morant M, Morant V M, Saito S, Bjarnholt N, Zagrobelny M, Jorgensen K, Osmani S, Simonsen H T, Perez R S, Heeswijck T B, Jorgensen B, Moller B L. Cyanogenic glycosides a case study for evolution and application of cytochromes P450. Phytochem Rev, 2006, 5: 309-329.
doi: 10.1007/s11101-006-9033-1 |
| [19] |
Hull A K, Vij R, Celenza J L. Arabidopsis cytochrome P450s that catalyze the first step of tryptophan-dependent indole-3-acetic acid biosynthesis. Proc Natl Acad Sci USA, 2000, 97: 2379-2384.
pmid: 10681464 |
| [20] |
Wittstock U, Halkier B A. Cytochrome 450 CYP79A2 from Arabidopsis thaliana L. catalyzes the conversion of l-phenylalanine to phenylacetaldoxime in the biosynthesis of benzylglucosinolate. J Biol Chem, 2000, 275: 14659-14666.
doi: 10.1074/jbc.275.19.14659 pmid: 10799553 |
| [21] |
Jorgensen K, Morant A V, Morant M, Jensen N B, Olsen C E, Kannangara R, Motawia M S, Moller B L, Bak S. Biosynthesis of the cyanogenic glucosides linamarin and lotaustralin in cassava isolation biochemical characterization and expression pattern of CYP71E7 the oxime-metabolizing cytochrome P450 enzyme. Plant Physiol, 2011, 155: 282-292.
doi: 10.1104/pp.110.164053 |
| [22] |
Luck K, Jia Q D, Huber M, Handrick V, Wong G K, Nelson D R, Chen F, Gershenzon J, Köllner T G. CYP79 P450 monooxygenases in gymnosperms: CYP79A118 is associated with the formation of taxiphyllin in Taxus baccata. Plant Mol Biol, 2017, 95: 169-180.
doi: 10.1007/s11103-017-0646-0 |
| [23] | Halkier B A, Hansen C H, Naur P, Mikkelsen M D, Wittstock U. The Role of Cytochromes P 450 in Biosynthesis and Evolution of Glucosinolates. Amsterdam: Elsevier Science Press, 2002. pp 223-248. |
| [24] |
Hu B, Jin J P, Guo A Y, Zhang H, Luo J C, Gao G. GSDS 2.0: anupgraded gene feature visualization server. Bioinformatics, 2015, 31: 1296-1297.
doi: 10.1093/bioinformatics/btu817 |
| [25] |
Wilkins M R, Gasteiger E, Bairoch A, Sanchez J C, Williams K L, Appel R D, Hochstrasser D F. Protein identification and analysis tools on the ExPASy server. Methods Mol Biol, 1999, 112: 531-552.
pmid: 10027275 |
| [26] | Bailey T L, Johnson J, Grant C E, Noble W S. The MEME suite. Nucleic Acids Res, 2015, 43: W39-W49. |
| [27] |
Wang Y P, Li J P, Paterson A H. MCScanX-transposed: detecting transposed gene duplications based on multiple collinearity scans. Bioinformatics, 2013, 29: 1458-1460.
doi: 10.1093/bioinformatics/btt150 |
| [28] |
Lynch M, Conery J S. The evolutionary fate and consequences of duplicate genes. Science, 2000, 290: 1151-1155.
doi: 10.1126/science.290.5494.1151 pmid: 11073452 |
| [29] | Lescot M, Dehais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, Rouze P, Rombauts S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res, 2002, 30: 325-327. |
| [30] |
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol, 2013, 30: 2725-2729.
doi: 10.1093/molbev/mst197 pmid: 24132122 |
| [31] |
Zhang J P, Qi Y N, Wang L M, Wang L L, Yan X C, Dang Z, Li W J, Zhao W, Pei X W, Li X M, Liu M, Tan M L, Wang L, Long Y, Wang J, Zhang X W, Dang Z H, Zheng H K, Liu T M. Genomic comparison and population diversity analysis provide insights into the domestication and improvement of flax. iScience, 2020, 23: 100967.
doi: 10.1016/j.isci.2020.100967 |
| [32] |
Huis R, Hawkins S, Neutelings G. Selection of reference genes for quantitative gene expression normalization in flax (Linum usitatissimum L.). BMC Plant Biol, 2010, 10: 71.
doi: 10.1186/1471-2229-10-71 |
| [33] |
Antonov J, Goldstein D R, Oberli A, Baltzer A, Pirotta M, Fleischmann A, Altermatt H J, Jaggi R. Reliable gene expression measurements from degraded RNA by quantitative real-time PCR depend on short amplicons and a proper normalization. Lab Invest, 2005, 85: 1040-1050.
pmid: 15951835 |
| [34] | 寇向龙, 徐美蓉, 张建平, 赵宾宾, 韩舜愈. 响应面法优化亚麻籽氢氰酸提取条件. 食品工业科技, 2016, 37(21): 201-204. |
| Kou X L, Xu M R, Zhang J P, Zhao B B, Han S Y. Optimization of extraction conditions for hydrocyanic acid in flaxseed by response surface methodology. Sci Technol Food Ind, 2016, 37(21): 201-204. (in Chinese with English abstract) | |
| [35] |
Mikkelsen M D, Hansen C H, Wittstock U, Halkier B A. Cytochrome 450 CYP79B2 from Arabidopsis catalyzes the conversion of tryptophan to indole-3-acetaldoxime, a precursor of indole glucosinolates and indole-3-acetic acid. J Biol Chem, 2000, 275: 33712-33717.
doi: 10.1074/jbc.M001667200 pmid: 10922360 |
| [36] | 李占省, 刘玉梅, 方智远, 杨丽梅, 庄木, 张扬勇, 吕红豪. 青花菜 P450 CYP79F1 全长克隆、表达及其与不同器官中莱菔硫烷含量的相关性分析. 中国农业科学, 2018, 51: 2357-2367. |
| Li Z S, Liu Y M, Fang Z Y, Yang L M, Zhuang M, Zhang Y Y, Lyu H H. Full length cloning, expression and correlation analysis of P450 CYP79F1 gene with sulforaphane content in different broccoli organs. Sci Agric Sin, 2018, 51: 2357-2367. (in Chinese with English abstract) | |
| [37] |
Ren R, Wang H F, Guo C C, Zhang N, Zeng L P, Chen Y M, Ma H, Qi J. Widespread whole genome duplications contribute to genome complexity and species diversity in angiosperms. Mol Plant, 2018, 11: 414-428.
doi: S1674-2052(18)30022-4 pmid: 29317285 |
| [38] |
Zhu Y, Wu N N, Song W L, Yin G J, Qin Y J, Yan Y M, Hu Y K. Soybean (Glycine max) expansin gene superfamily origins: segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol, 2014, 14: 93.
doi: 10.1186/1471-2229-14-93 pmid: 24720629 |
| [39] | 华静静, 陈晓静. 植物体中WD40蛋白的研究进展. 黑龙江农业科学, 2015, (5): 153-156. |
| Hua J J, Chen X J. Progress of WD40 proteins in plant. Heilongjiang Agric Sci, 2015, (5): 153-156 (in Chinese with English abstract). | |
| [40] |
Mehlgarten C, Jablonowski D, Wrackmeyer U, Tschitschmann S, Sondermann D, Jäger G, Gong Z, Byström A S, Schaffrath R, Breunig K D. Elongator function in tRNA wobble uridine modification is conserved between yeast and plants. Mol Microbiol, 2010, 76: 1082-1094.
doi: 10.1111/j.1365-2958.2010.07163.x pmid: 20398216 |
| [41] | 杨琳, 王宇, 杨剑飞, 李玉花. 花青素积累相关负调控因子的研究进展. 园艺学报, 2014, 41: 1873-1884. |
| Yang L, Wang Y, Yang J F, Li Y H. Research advances on negative regulators of anthocyanin accumulation. Acta Hortic Sin, 2014, 41: 1873-1884. (in Chinese with English abstract) |
| [1] | 孙全喜, 苑翠玲, 牟艺菲, 闫彩霞, 赵小波, 王娟, 王奇, 孙慧, 李春娟, 单世华. 花生SWEET基因全基因组鉴定及表达分析[J]. 作物学报, 2023, 49(4): 938-954. |
| [2] | 潘洁明, 田绍锐, 梁艳兰, 朱宇林, 周定港, 阙友雄, 凌辉, 黄宁. 甘蔗PIN-LIKES基因家族的鉴定与表达分析[J]. 作物学报, 2023, 49(2): 414-425. |
| [3] | 濮雪, 王凯彤, 张宁, 司怀军. 马铃薯StMAPKK4基因表达分析及互作蛋白筛选与鉴定[J]. 作物学报, 2023, 49(1): 36-45. |
| [4] | 王沙沙, 黄超, 汪庆昌, 晁岳恩, 陈锋, 孙建国, 宋晓. 小麦籽粒大小相关基因TaGS2克隆及功能分析[J]. 作物学报, 2022, 48(8): 1926-1937. |
| [5] | 陈璐, 周淑倩, 李永新, 陈刚, 陆国权, 杨虎清. 甘薯解偶联蛋白基因家族鉴定与表达分析[J]. 作物学报, 2022, 48(7): 1683-1696. |
| [6] | 陈松余, 丁一娟, 孙峻溟, 黄登文, 杨楠, 代雨涵, 万华方, 钱伟. 甘蓝型油菜BnCNGC基因家族鉴定及其在核盘菌侵染和PEG处理下的表达特性分析[J]. 作物学报, 2022, 48(6): 1357-1371. |
| [7] | 晋敏姗, 曲瑞芳, 李红英, 韩彦卿, 马芳芳, 韩渊怀, 邢国芳. 谷子糖转运蛋白基因SiSTPs的鉴定及其参与谷子抗逆胁迫响应的研究[J]. 作物学报, 2022, 48(4): 825-839. |
| [8] | 巫燕飞, 胡琴, 周棋, 杜雪竹, 盛锋. 水稻延伸因子复合体家族基因鉴定及非生物胁迫诱导表达模式分析[J]. 作物学报, 2022, 48(3): 644-655. |
| [9] | 靳容, 蒋薇, 刘明, 赵鹏, 张强强, 李铁鑫, 王丹凤, 范文静, 张爱君, 唐忠厚. 甘薯Dof基因家族挖掘及表达分析[J]. 作物学报, 2022, 48(3): 608-623. |
| [10] | 林焕泰, 张天杰, 史梦婷, 郭燕芳, 高三基, 王锦达. 割手密萜烯合成酶(TPS)基因家族分析及其在生物胁迫下的表达分析[J]. 作物学报, 2022, 48(12): 3029-3044. |
| [11] | 马文婧, 刘震, 李志涛, 朱金勇, 李泓阳, 陈丽敏, 史田斌, 张俊莲, 刘玉汇. 马铃薯BBX基因家族的全基因组鉴定及表达分析[J]. 作物学报, 2022, 48(11): 2797-2812. |
| [12] | 贾小霞, 齐恩芳, 马胜, 黄伟, 郑永伟, 白永杰, 文国宏. 马铃薯PYL基因家族的全基因组鉴定及表达分析[J]. 作物学报, 2022, 48(10): 2533-2545. |
| [13] | 余慧芳, 张卫娜, 康益晨, 范艳玲, 杨昕宇, 石铭福, 张茹艳, 张俊莲, 秦舒浩. 马铃薯CrRLK1Ls基因家族的鉴定及响应晚疫病菌信号的表达分析[J]. 作物学报, 2022, 48(1): 249-258. |
| [14] | 荐红举, 尚丽娜, 金中辉, 丁艺, 李燕, 王季春, 胡柏耿, Vadim Khassanov, 吕典秋. 马铃薯PIF家族成员鉴定及其对高温胁迫的响应分析[J]. 作物学报, 2022, 48(1): 86-98. |
| [15] | 谢琴琴, 左同鸿, 胡燈科, 刘倩莹, 张以忠, 张贺翠, 曾文艺, 袁崇墨, 朱利泉. 甘蓝自交不亲和相关基因BoPUB9的克隆及表达分析[J]. 作物学报, 2022, 48(1): 108-120. |
|