Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2022, Vol. 48 ›› Issue (5): 1119-1128.doi: 10.3724/SP.J.1006.2022.12022
• CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS • Previous Articles Next Articles
YANG De-Wei1,2( ), WANG Xun1(), ZHENG Xing-Xing1, XIANG Xin-Quan1, CUI Hai-Tao1, LI Sheng-Ping1,*(), TANG Ding-Zhong1,*()
), WANG Xun1(), ZHENG Xing-Xing1, XIANG Xin-Quan1, CUI Hai-Tao1, LI Sheng-Ping1,*(), TANG Ding-Zhong1,*()
| [1] | Zhang N, Luo J, Rossman A Y, Aoki T, Chuma I, Crous P W, Dean R, de Vries R P, Donofrio N, Hyde K D, Lebrun M H, Talbot N J, Tharreau D, Tosa Y, Valent B, Wang Z H, Xu J R. Generic names in Magnaporthales. IMA Fung, 2016, 7:155-159. |
| [2] |
Li W T, Chern M S, Yin J J, Wang J, Chen X W. Recent advances in broad-spectrum resistance to the rice blast disease. Curr Opin Plant Biol, 2019, 50:114-120.
doi: 10.1016/j.pbi.2019.03.015 |
| [3] |
Deng Y W, Zhai K R, Xie Z, Yang D Y, Zhu X D, Liu J Z, Wang X, Qin P, Yang Y Z, Zhang G M, Li Q, Zhang J F, Wu S Q, Milazzo J, Mao B Z, Wang E T, Xie H A, Tharreau D, He Z H. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science, 2017, 355:962-965.
doi: 10.1126/science.aai8898 |
| [4] |
Yang D W, Li S P, Lu L, Fang J B, Wang W, Cui H T, Tang D Z. Identification and application of the Pigm-1 gene in rice disease-resistance breeding. Plant Biol, 2020, 22:1022-1029.
doi: 10.1111/plb.v22.6 |
| [5] |
Zhou B, Qu S H, Liu G F, Dolan M, Sakai H, Lu G D, Bellizzi M, Wang G L. The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea. Mol Plant Microbe Interact, 2006, 19:1216-1228.
doi: 10.1094/MPMI-19-1216 |
| [6] |
Zhu X Y, Chen S, Yang J Y, Zhou S C, Zeng L X, Han J L, Su J, Wang, Pan Q H. The identification of Pi50(t), a new member of the rice blast resistance Pi2/Pi9 multigene family. Theor Appl Genet, 2012, 124:1295-1304.
doi: 10.1007/s00122-012-1787-9 |
| [7] |
Jiang N, Li Z Q, Wu J, Wang Y, Wu L Q, Wang S H, Wang D, Wen T, Liang Y, Sun P Y, Liu J L, Dai L Y, Wang Z L, Wang C, Luo M Z, Liu X L, Wang G L. Molecular mapping of the Pi2/9 allelic gene Pi2-2 conferring broad-spectrum resistance to Magnaporthe oryzae in the rice cultivar Jefferson. Rice, 2012, 5:29.
doi: 10.1186/1939-8433-5-29 pmid: 27234247 |
| [8] |
Su J, Wang W J, Han J L, Chen S, Wang C Y, Zeng L X, Feng A Q, Yang J Y, Zhou B, Zhu X Y. Functional divergence of duplicated genes results in a novel blast resistance gene Pi50 at the Pi2/9 locus. Theor Appl Genet, 2015, 128:2213-2225.
doi: 10.1007/s00122-015-2579-9 |
| [9] |
Deng Y W, Zhai K R, Xie Z, Yang D Y, Zhu X D, Liu J Z, Wang X, Qin P, Yang Y Z, Zhang G M, Li Q, Zhang J F, Wu S Q, Milazzo J, Mao B Z, Wang E T, Xie H A, Tharreau D, He Z H. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science, 2017, 355:962-965.
doi: 10.1126/science.aai8898 |
| [10] | Dodds P N, Rathjen J P. Plant immunity: towards an integrated view of plant-pathogen interactions. Nat Rev Genet, 2010, 11:539-548. |
| [11] |
Wang W, Feng B M, Zhou J M, Tang D Z. Plant immune signaling: advancing on two frontiers. J Integr Plant Biol, 2020, 62:2-24.
doi: 10.1111/jipb.v62.1 |
| [12] | 杨德卫, 李生平, 崔海涛, 邹声浩, 王伟. 寄主植物与病原菌免疫反应的分子遗传基础. 遗传, 2020, 42:278-286. |
| Yang D W, Li S P, Cui H T, Zou S H, Wang W. Molecular genetic mechanisms of interaction between host plants and pathogens. Hereditas(Beijing), 2020, 42:278-286 (in Chinese with English abstract). | |
| [13] |
Jones J D G, Dangl J L. The plant immune system. Nature, 2006, 444:323-329.
doi: 10.1038/nature05286 |
| [14] | Yuan M, Jiang Z, Bi G, Nomura K, Liu M, Wang Y, Cai B, Zhou J M, He S Y, Xin X F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature, 2021, 592:105-109. |
| [15] | Ngou B P M, Ahn H K, Ding P T, Jones J D G. Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature, 2021, 592:110-115. |
| [16] |
Li W T, Chern M S, Yin J J, Wang J, Chen X W. Recent advances in broad-spectrum resistance to the rice blast disease. Curr Opin Plant Biol, 2019, 50:114-120
doi: 10.1016/j.pbi.2019.03.015 |
| [17] | Hayafune M, Berisio R, Marchetti R, Silipo A, Kayama M, Desaki Y, Arima S, Squeglia F, Ruggiero A, Tokuyasu K, Molinaro A, Kaku H, Shibuya N. Chitin-induced activation of immune signaling by the rice receptor CEBiP relies on a unique sandwich-type dimerization. Proc Natl Acad Sci USA, 2014, 111:404-413. |
| [18] |
Yamaguchi K, Yamada K, Ishikawa K, Yoshimura S, Hayashi N, Uchihashi K, Ishihama N, Kishi-Kaboshi M, Takahashi A, Tsuge S, Ochiai H, Tada Y, Shimamoto K, Yoshioka H, Kawasaki T. A receptor-like cytoplasmic kinase targeted by a plant pathogen effector is directly phosphorylated by the chitin receptor and mediates rice immunity. Cell Host Microb, 2013, 13:347-357.
doi: 10.1016/j.chom.2013.02.007 |
| [19] |
Yamada K, Yamaguchi K, Yoshimura S, Terauchi A, Kawasaki T. Conservation of chitin-induced MAPK signaling pathways in rice and Arabidopsis. Plant Cell Physiol, 2017, 58:993-1002.
doi: 10.1093/pcp/pcx042 pmid: 28371870 |
| [20] |
Wang C, Wang G, Zhang C, Zhu P K, Dai H L, Yu N, He Z H, Xu L, Wang E T. OsCERK1-mediated chitin perception and immune signaling requires receptor-like cytoplasmic kinase 185 to activate an MAPK cascade in rice. Mol Plant, 2017, 10:619-633.
doi: 10.1016/j.molp.2017.01.006 |
| [21] |
Pennisi E. Armed and dangerous. Science, 2010, 327:804-805.
doi: 10.1126/science.327.5967.804 |
| [22] |
Mine A, Seyerth C, Kracher B, Berens M L, Becker D, Tsuda K. The defense phytohormone signaling network enables rapid, high-amplitude transcriptional reprogramming during eector- triggered immunity. Plant Cell, 2018, 30:1199-1219.
doi: 10.1105/tpc.17.00970 |
| [23] |
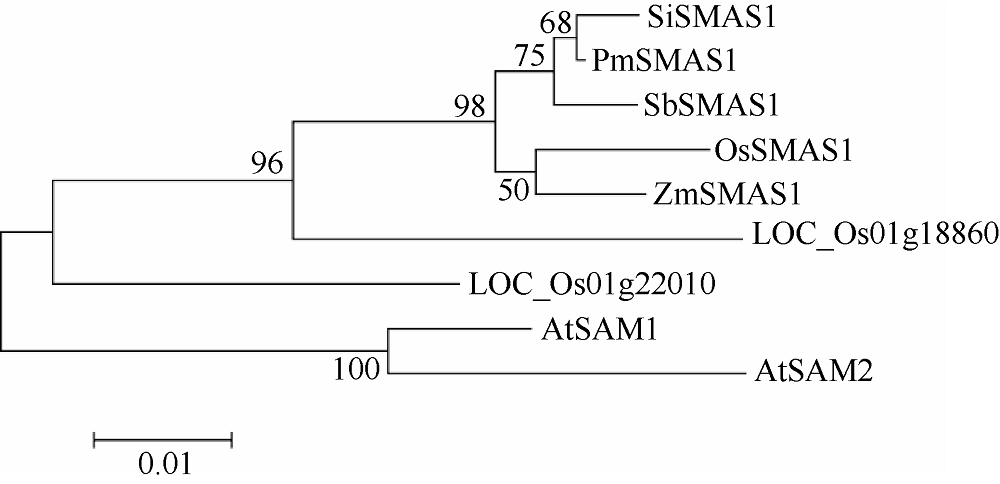
Meng J J, Wang L S, Wang J Y, Zhao X W, Cheng J K, Yu W X, Jin D, Li Q, Gong Z Z. Methionine adenosyltransferase4 mediates DNA and histone methylation. Plant Physiol, 2018, 177:652-670.
doi: 10.1104/pp.18.00183 |
| [24] |
Yan, X J, Ma L, Pang H Y, Wang P, Lei L, Cheng Y X, Cheng J K, Guo Y, Li Q Z. Methionine synthase1 is involved in chromatin silencing by maintaining dna and histone methylation. Plant Physiol, 2019, 181:249-261.
doi: 10.1104/pp.19.00528 |
| [25] |
Chen Y, Xu Y Y, Luo W, Li W X, Chen N, Zhang D J, Chong K. The F-box protein OsFBK12 targets OsSAMS1 for degradation and affects pleiotropic phenotypes, including leaf senescence, in rice. Plant Physiol, 2013, 163:1673-1685.
doi: 10.1104/pp.113.224527 |
| [26] |
Bustin S A, Benes V, Garson J A, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl M W, Shipley G L, Vandesompe J, Wittwer C T. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem, 2009, 55:611-622.
doi: 10.1373/clinchem.2008.112797 |
| [27] |
Park C H, Chen S B, Shirsekar G, Zhou B, Khang C H, Songkumarn P, Afzal A J, Ning Y S, Wang R S, Bellizzi M, Valent B, Wang G L. The Magnaporthe oryzae effector AvrPiz-t targets the RING E3 ubiquitin ligase APIP6 to suppress pathogen-associated molecular pattern-triggered immunity in rice. Plant Cell, 2012, 24:4748-4762.
doi: 10.1105/tpc.112.105429 |
| [28] |
Yang D W, Cheng C P, Zheng X H, Ye X F, Ye N, Huang F H. Identification and fine mapping of a major QTL, qHD19, that plays pleiotropic roles in regulating the heading date in rice. Mol Breed, 2020, 40:30.
doi: 10.1007/s11032-020-1109-x |
| [29] |
Schwessinger B, Ronald P C. Plant innate immunity: perception of conserved microbial signatures. Annu Rev Plant Biol, 2012, 63:451-482.
doi: 10.1146/annurev-arplant-042811-105518 pmid: 22404464 |
| [30] |
Yang C, Li W, Cao J D, Meng F W, Yu Y Q, Huang J K, Jiang L, Liu M X, Zhang Z G, Chen X W, Miyamoto K, Yamane H, Zhang J S, Chen S Y, Liu J. Activation of ethylene signaling pathways enhances disease resistance by regulating ROS and phytoalexin production in rice. Plant J, 2017, 89:338-353.
doi: 10.1111/tpj.13388 |
| [31] |
Mao D, Feng Y, Jian L, Poel B V, Tan D, Li J L, Liu Y Q, Li X S, Dong M Q, Chen L B, Li D P, Luan S. FERONIA receptor kinase interacts with S-adenosylmethionine synthetase and suppresses S-adenosylmethionine production and ethylene biosynthesis in Arabidopsis. Plant Cell Environ, 2016, 38:2566-2574.
doi: 10.1111/pce.12570 |
| [32] |
Ji D C, Cui X M, Qin G Z, Chen T, Tian S P. SlFERL interacts with S-adenosylmethionine synthetase to regulate fruit ripening. Plant Physiol, 2020, 184:2168-2181.
doi: 10.1104/pp.20.01203 |
| [33] |
Li W X, Han Y Y, Tao F, Chong K. Knockdown of SAMS genes encoding S-adenosyl-L-methionine synthetases causes methylation alterations of DNAs and histones and leads to late flowering in rice. J Plant Physiol, 2011, 168:1837-1843.
doi: 10.1016/j.jplph.2011.05.020 |
| [34] |
Iwai T, Miyasaka A, Seo S, Ohashi Y. Contribution of ethylene biosynthesis for resistance to blast fungus infection in young rice plants. Plant Physiol, 2006, 142:1202-1215.
doi: 10.1104/pp.106.085258 |
| [35] |
Tintor N, Ross A, Kanehara K, Yamada K, Fan L, Kemmerling B, Nürnberger T, Tsuda K, Saijo Y. Layered pattern receptor signaling via ethylene and endogenous elicitor peptides during Arabidopsis immunity to bacterial infection. Proc Natl Acad Sci USA, 2013, 110:6211-6216.
doi: 10.1073/pnas.1216780110 |
| [36] |
Helliwell E E, Wang Q, Yang Y N. Transgenic rice with inducible ethylene production exhibits broad-spectrum disease resistance to the fungal pathogens Magnaporthe oryzae and Rhizoctonia solani. Plant Biotechnol J, 2013, 11:33-42.
doi: 10.1111/pbi.12004 pmid: 23031077 |
| [37] |
Singh M P, Lee F N, Counce P A, Gibbons J H. Mediation of partial resistance to rice blast through anaerobic induction of ethylene. Phytopathology, 2004, 94:819-825.
doi: 10.1094/PHYTO.2004.94.8.819 |
| [38] | Seo Y S, Chern M, Bartley L E, Han M, Jung K H, Lee I, Walia H, Richter T, Xu X, Cao P, Bai W, Ramanan R, Amonpant F, Arul L, Canlas P E, Ruan R, Park C J, Chen X, Hwang S, Jeon J S, Ronald P C. Towards establishment of a rice stress response interactome. PLoS Genet, 2011, 7:e1002020. |
| [39] |
Gong B, Li X VandenLangenberg K M, Wen D, Sun S S, Wei M, Li Y, Yang F J, Shi Q H, Wang X F. Overexpression of S-adenosyl-L-methionine synthetase increased tomato tolerance to alkali stress through polyamine metabolism. Plant Biotechnol J, 2014, 12:694-708.
doi: 10.1111/pbi.2014.12.issue-6 |
| [40] |
Mao D D, Yu F, Li J, Van de Poel B, Tan D, Li J L, Liu Y Q, Li X S, Dong M Q, Chen L B, Li D P, Luan S. FERONIA receptor kinase interacts with S-denosylmethionine synthetase and suppresses S-denosylmethionine production and ethylene biosynthesis in Arabidopsis. Plant Cell Environ, 2015, 38:2566-2574.
doi: 10.1111/pce.12570 |
| [41] |
Chen Y, Zou T, McCormick S. S-adenosylmethionine synthetase 3 is important for pollen tube growth. Plant Physiol, 2016, 172:244-253.
doi: 10.1104/pp.16.00774 |
| [42] |
Shen B, Li C, Tarczynski M C. High free-methionine and decreased lignin content result from a mutation in the Arabidopsis S-adenosyl-L-methionine synthetase 3 gene. Plant J, 2002, 29:371-380.
pmid: 11844113 |
| [43] |
Li W X, Han Y Y, Tao F, Chong K. Knockdown of SAMS genes encoding S-adenosyl-l-methionine synthetases causes methylation alterations of DNAs and histones and leads to late flowering in rice. J Plant Physiol, 2011, 168:1837-1843.
doi: 10.1016/j.jplph.2011.05.020 |
| [1] | TIAN Tian, CHEN Li-Juan, HE Hua-Qin. Identification of rice blast resistance candidate genes based on integrating Meta-QTL and RNA-seq analysis [J]. Acta Agronomica Sinica, 2022, 48(6): 1372-1388. |
| [2] | ZHENG Chong-Ke, ZHOU Guan-Hua, NIU Shu-Lin, HE Ya-Nan, SUN wei, XIE Xian-Zhi. Phenotypic characterization and gene mapping of an early senescence leaf H5(esl-H5) mutant in rice (Oryza sativa L.) [J]. Acta Agronomica Sinica, 2022, 48(6): 1389-1400. |
| [3] | ZHOU Wen-Qi, QIANG Xiao-Xia, WANG Sen, JIANG Jing-Wen, WEI Wan-Rong. Mechanism of drought and salt tolerance of OsLPL2/PIR gene in rice [J]. Acta Agronomica Sinica, 2022, 48(6): 1401-1415. |
| [4] | ZHENG Xiao-Long, ZHOU Jing-Qing, BAI Yang, SHAO Ya-Fang, ZHANG Lin-Ping, HU Pei-Song, WEI Xiang-Jin. Difference and molecular mechanism of soluble sugar metabolism and quality of different rice panicle in japonica rice [J]. Acta Agronomica Sinica, 2022, 48(6): 1425-1436. |
| [5] | YAN Jia-Qian, GU Yi-Biao, XUE Zhang-Yi, ZHOU Tian-Yang, GE Qian-Qian, ZHANG Hao, LIU Li-Jun, WANG Zhi-Qin, GU Jun-Fei, YANG Jian-Chang, ZHOU Zhen-Ling, XU Da-Yong. Different responses of rice cultivars to salt stress and the underlying mechanisms [J]. Acta Agronomica Sinica, 2022, 48(6): 1463-1475. |
| [6] | YANG Jian-Chang, LI Chao-Qing, JIANG Yi. Contents and compositions of amino acids in rice grains and their regulation: a review [J]. Acta Agronomica Sinica, 2022, 48(5): 1037-1050. |
| [7] | DENG Zhao, JIANG Nan, FU Chen-Jian, YAN Tian-Zhe, FU Xing-Xue, HU Xiao-Chun, QIN Peng, LIU Shan-Shan, WANG Kai, YANG Yuan-Zhu. Analysis of blast resistance genes in Longliangyou and Jingliangyou hybrid rice varieties [J]. Acta Agronomica Sinica, 2022, 48(5): 1071-1080. |
| [8] | ZHU Zheng, WANG Tian-Xing-Zi, CHEN Yue, LIU Yu-Qing, YAN Gao-Wei, XU Shan, MA Jin-Jiao, DOU Shi-Juan, LI Li-Yun, LIU Guo-Zhen. Rice transcription factor WRKY68 plays a positive role in Xa21-mediated resistance to Xanthomonas oryzae pv. oryzae [J]. Acta Agronomica Sinica, 2022, 48(5): 1129-1140. |
| [9] | WANG Xiao-Lei, LI Wei-Xing, OU-YANG Lin-Juan, XU Jie, CHEN Xiao-Rong, BIAN Jian-Min, HU Li-Fang, PENG Xiao-Song, HE Xiao-Peng, FU Jun-Ru, ZHOU Da-Hu, HE Hao-Hua, SUN Xiao-Tang, ZHU Chang-Lan. QTL mapping for plant architecture in rice based on chromosome segment substitution lines [J]. Acta Agronomica Sinica, 2022, 48(5): 1141-1151. |
| [10] | WANG Ze, ZHOU Qin-Yang, LIU Cong, MU Yue, GUO Wei, DING Yan-Feng, NINOMIYA Seishi. Estimation and evaluation of paddy rice canopy characteristics based on images from UAV and ground camera [J]. Acta Agronomica Sinica, 2022, 48(5): 1248-1261. |
| [11] | KE Jian, CHEN Ting-Ting, WU Zhou, ZHU Tie-Zhong, SUN Jie, HE Hai-Bing, YOU Cui-Cui, ZHU De-Quan, WU Li-Quan. Suitable varieties and high-yielding population characteristics of late season rice in the northern margin area of double-cropping rice along the Yangtze River [J]. Acta Agronomica Sinica, 2022, 48(4): 1005-1016. |
| [12] | CHEN Yue, SUN Ming-Zhe, JIA Bo-Wei, LENG Yue, SUN Xiao-Li. Research progress regarding the function and mechanism of rice AP2/ERF transcription factor in stress response [J]. Acta Agronomica Sinica, 2022, 48(4): 781-790. |
| [13] | WANG Lyu, CUI Yue-Zhen, WU Yu-Hong, HAO Xing-Shun, ZHANG Chun-Hui, WANG Jun-Yi, LIU Yi-Xin, LI Xiao-Gang, QIN Yu-Hang. Effects of rice stalks mulching combined with green manure (Astragalus smicus L.) incorporated into soil and reducing nitrogen fertilizer rate on rice yield and soil fertility [J]. Acta Agronomica Sinica, 2022, 48(4): 952-961. |
| [14] | QIN Qin, TAO You-Feng, HUANG Bang-Chao, LI Hui, GAO Yun-Tian, ZHONG Xiao-Yuan, ZHOU Zhong-Lin, ZHU Li, LEI Xiao-Long, FENG Sheng-Qiang, WANG Xu, REN Wan-Jun. Characteristics of panicle stem growth and flowering period of the parents of hybrid rice in machine-transplanted seed production [J]. Acta Agronomica Sinica, 2022, 48(4): 988-1004. |
| [15] | WU Yan-Fei, HU Qin, ZHOU Qi, DU Xue-Zhu, SHENG Feng. Genome-wide identification and expression analysis of Elongator complex family genes in response to abiotic stresses in rice [J]. Acta Agronomica Sinica, 2022, 48(3): 644-655. |
|
||