欢迎访问作物学报,今天是
作物学报 ›› 2025, Vol. 51 ›› Issue (1): 247-259.doi: 10.3724/SP.J.1006.2025.44044
宋倩娜1,2( ), 宋慧洋1, 李京昊1, 段永红1, 梅超1,2, 冯瑞云1,2,*()
), 宋慧洋1, 李京昊1, 段永红1, 梅超1,2, 冯瑞云1,2,*()
SONG Qian-Na1,2(), SONG Hui-Yang1, LI Jing-Hao1, DUAN Yong-Hong1, MEI Chao1,2, FENG Rui-Yun1,2,*()
摘要:
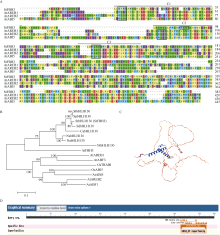
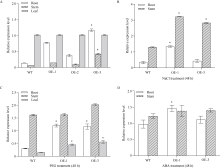
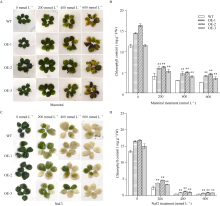
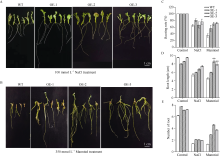
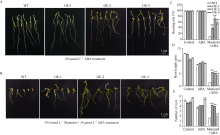
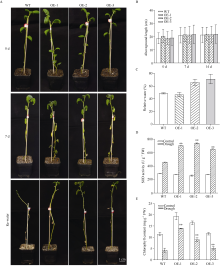
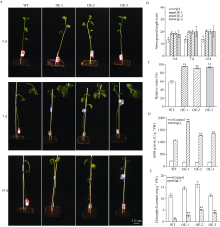
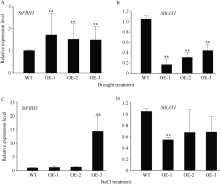
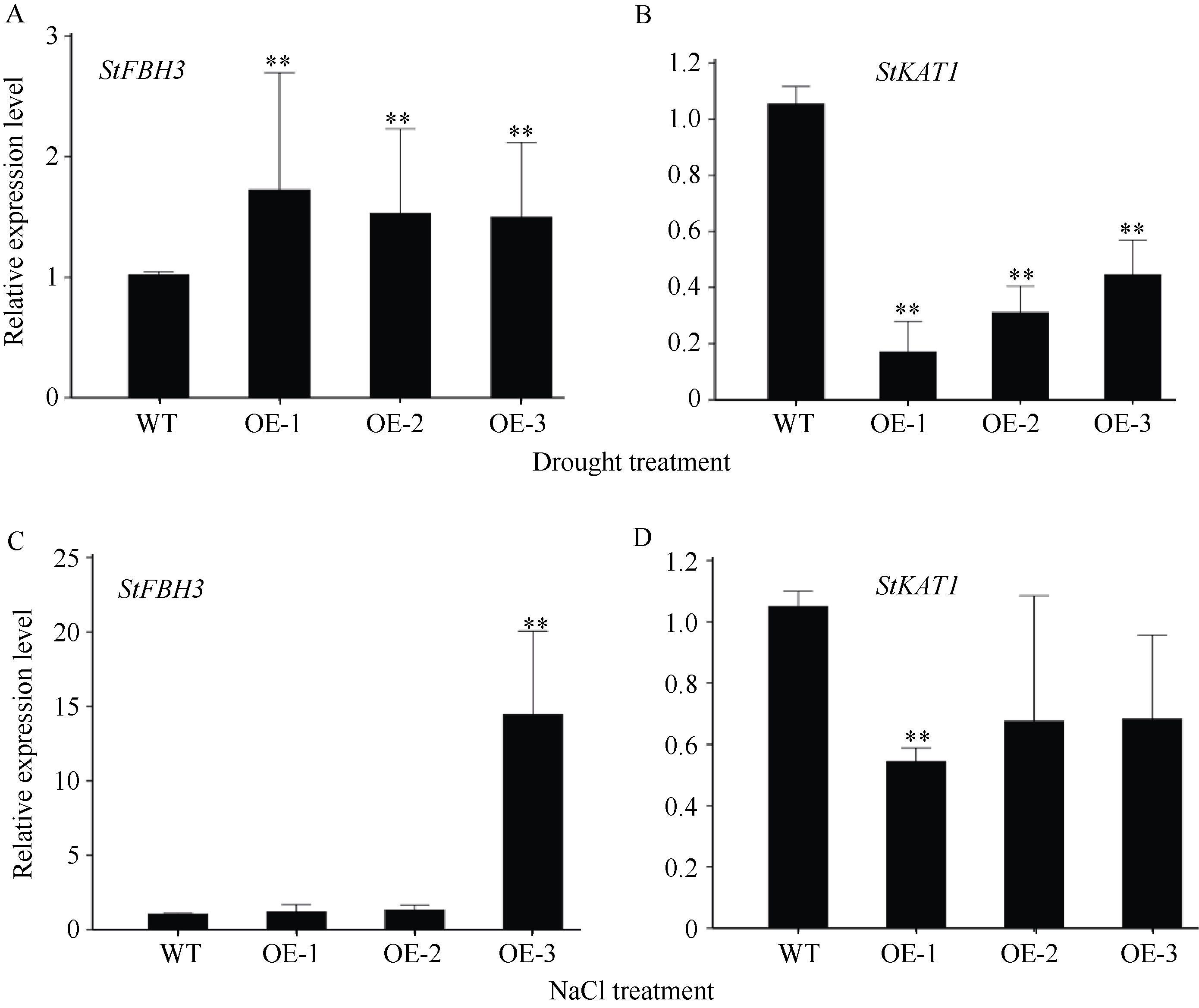
bHLH (basic helix-loop-helix)作为植物界第二大类转录因子, 在植物应对环境胁迫响应中起着重要的调控作用。探究马铃薯(Solanum tuberosum L.) bHLH家族基因功能将为马铃薯改良和育种提供一定的理论依据。本研究克隆了马铃薯StFBH3基因(Gene ID: 102582309), 利用qPCR技术分析了不同逆境胁迫下StFBH3基因表达模式, 结果表明, StFBH3基因在马铃薯根和叶中的表达量较高, 且该基因的表达受渗透、高盐和脱落酸(abscisic acid, ABA)诱导; 以过表达StFBH3基因的马铃薯试管苗为材料, 在分别含有不同浓度甘露醇(mannitol)、NaCl和ABA的MS培养基中, 过表达StFBH3马铃薯株系的叶绿素含量显著高于野生型, 根长显著长于野生型。在干旱和高盐处理下, 土壤栽培的过表达马铃薯株系较野生型表现较强的耐受性, 叶片相对含水量、叶绿素含量和过氧化物歧化酶(SOD)活性显著高于野生型。qPCR分析发现, 干旱和盐胁迫处理下相关基因(KAT1)的表达量在过表达马铃薯株系中较野生型显著降低。以上结果表明, StFBH3基因可能在马铃薯对渗透、干旱和高盐等胁迫响应中起正向调控作用。本研究也为StFBH3基因在马铃薯中的生物学功能深入理解提供一定的参考依据。
| [1] | 张冠初, 张智猛, 慈敦伟, 丁红, 杨吉顺, 史晓龙, 田家明, 戴良香. 干旱和盐胁迫对花生渗透调节和抗氧化酶活性的影响. 华北农学报, 2018, 33(3): 176-181. |
| Zhang G C, Zhang Z M, Ci D W, Ding H, Yang J S, Shi X L, Tian J M, Dai L X. Effects of drought and salt stress on osmotic regulator and antioxidase activities. Acta Agric Boreali-Sin, 2018, 33(3): 176-181 (in Chinese with English abstract). | |
| [2] | Munns R, Tester M. Mechanisms of salinity tolerance. Annu Rev Plant Biol, 2008, 59: 651-681. |
| [3] | Suzuki N, Koussevitzky S, Mittler R, Miller G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ, 2012, 35: 259-270. |
| [4] | 仇焕广, 李新海, 余嘉玲. 中国玉米产业: 发展趋势与政策建议. 农业经济问题, 2021, 42(7): 4-16. |
| Qiu H G, Li X H, Yu J L. China maize industry: development trends and policy suggestions. Issues Agric Econ, 2021, 42(7): 4-16 (in Chinese with English abstract). | |
| [5] | 张雪莹, 刘欣. 转录因子与叶片发育的研究进展. 植物生理学报, 2022, 58: 91-100. |
| Zhang X Y, Liu X. Research progress of transcription factors and leaf development. Plant Physiol J, 2022, 58: 91-100 (in Chinese with English abstract). | |
| [6] | Balazadeh S, Riaño-Pachón D M, Mueller-Roeber B. Transcription factors regulating leaf senescence in Arabidopsis thaliana. Plant Cell, 2008, 10: 63-75. |
| [7] | Ng D W, Abeysinghe J K, Kamali M. Regulating the regulators: the control of transcription factors in plant defense signaling. Int J Mol Sci, 2018, 19: 3737. |
| [8] | Kong Q, Pattanaik S, Feller A, Werkman J R, Chai C L, Wang Y Q, Grotewold E, Yuan L. Regulatory switch enforced by basic helix-loop-helix and ACT-domain mediated dimerizations of the maize transcription factor R. Proc Natl Acad Sci USA, 2012, 109: E2091-E2097. |
| [9] | Pires N, Dolan L. Origin and diversification of basic-helix-loop- helix proteins in plants. Mol Biol Evol, 2010, 27: 862-874. |
| [10] | Filiz E, Vatansever R, Ozyigit I I. Dissecting a co-expression network of basic helix-loop-helix (bHLH) genes from phosphate (Pi)-starved soybean (Glycine max). Plant Gene, 2017, 9: 19-25. |
| [11] | Murre C, Bain G, van Dijk M A, Engel I, Furnari B A, Massari M E, Matthews J R, Quong M W, Rivera R R, Stuiver M H. Structure and function of helix-loop-helix proteins. Biochim Biophys Acta, 1994, 1218: 129-135. |
| [12] | Zhang Z S, Chen J, Liang C L, Liu F, Hou X L, Zou X X. Genome-wide identification and characterization of the bHLH transcription factor family in pepper (Capsicum annuum L.). Front Genet, 2020, 11: 570156. |
| [13] | 杨梦婷, 张春, 王作平, 邹华文, 吴忠义. 玉米ZmbHLH161基因的克隆及功能研究. 作物学报, 2020, 46: 2008-2016. |
| Yang M T, Zhang C, Wang Z P, Zou H W, Wu Z Y. Cloning and functional analysis of ZmbHLH161gene in maize. Acta Agron Sin, 2020, 46: 2008-2016 (in Chinese with English abstract). | |
| [14] | Li X X, Duan X P, Jiang H X, Sun Y J, Tang Y P, Yuan Z, Guo J K, Liang W Q, Chen L, Yin J Y, Ma H, Wang J, Zhang D B. Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Cell, 2006, 141: 1167-1184. |
| [15] | Tian S Y, Li L J, Wei M, Yang F J. Genome-wide analysis of basic helix-loop-helix superfamily members related to anthocyanin biosynthesis in eggplant (Solanum melongena L.). PeerJ, 2019, 7: e7768. |
| [16] | 冯建英, 李立芹, 鲁黎明. 马铃薯bHLH转录因子家族全基因组鉴定与表达分析. 生物技术通报, 2022, 38(2): 21-33. |
| Feng J Y, Li L Q, Lu L M. Genome-wide identification and expression analysis of the bHLH transcription factor family in Solanum tuberosum. Plant Cell, 2022, 38(2): 21-33 (in Chinese with English abstract). | |
| [17] | Daie J. Annual review of plant physiology and plant molecular biology. Soil Sci, 1992, 154: 508. |
| [18] | Le Hir R, Castelain M, Chakraborti D, Moritz T, Dinant S, Bellini C. AtbHLH68 transcription factor contributes to the regulation of ABA homeostasis and drought stress tolerance in Arabidopsis thaliana. Plant Cell, 2017, 160: 312-327. |
| [19] | Zhao Q, Fan Z H, Qiu L N, Che Q Q, Wang T, Li Y Y, Wang Y Z. MdbHLH130, an apple bHLH transcription factor, confers water stress resistance by regulating stomatal closure and ROS homeostasis in transgenic tobacco. Front Plant Sci, 2020, 11: 543696. |
| [20] | Zhou J, Li F, Wang J L, Ma Y, Chong K, Xu Y Y. Basic helix-loop-helix transcription factor from wild rice (OrbHLH2) improves tolerance to salt- and osmotic stress in Arabidopsis. Plant Cell, 2009, 166: 1296-1306. |
| [21] | Busk P K, Pagès M. Regulation of abscisic acid-induced transcription. Plant Mol Biol, 1998, 37: 425-435. |
| [22] | Suzuki M, Ketterling M G, Li Q B, McCarty D R. Viviparous1 alters global gene expression patterns through regulation of abscisic acid signaling. Plant Physiol, 2003, 132: 1664-1677. |
| [23] | Fujita Y, Fujita M, Satoh R, Maruyama K, Parvez M M, Seki M, Hiratsu K, Ohme-Takagi M, Shinozaki K, Yamaguchi-Shinozaki K. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell, 2005, 17: 3470-3488. |
| [24] | Hobo T, Kowyama Y, Hattori T. A bZIP factor, TRAB1, interacts with VP1 and mediates abscisic acid-induced transcription. Proc Natl Acad Sci USA, 1999, 96: 15348-15353. |
| [25] | Casaretto J, Ho T H D. The transcription factors HvABI5 and HvVP1 are required for the abscisic acid induction of gene expression in barley aleurone cells. Plant Cell, 2003, 15: 271-284. |
| [26] | Furihata T, Maruyama K, Fujita Y, Umezawa T, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K. Abscisic acid-dependent multisite phosphorylation regulates the activity of a transcription activator AREB1. Proc Natl Acad Sci USA, 2006, 103: 1988-1993. |
| [27] | Takahashi Y, Kinoshita T, Matsumoto M, Shimazaki K. Inhibition of the Arabidopsis bHLH transcription factor by monomerization through abscisic acid-induced phosphorylation. Plant J, 2016, 87: 559-567. |
| [28] | Halterman D, Guenthner J, Collinge S, Butler N, Douches D. Biotech potatoes in the 21st century: 20 years since the first biotech potato. Am J Potato Res, 2016, 93: 1-20. |
| [29] | Suttle J. Symposium introduction:enhancing the nutritional value of potato tubers. Am J Potato Res, 2008, 85: 266. |
| [30] | Liu F L, Shahnazari A, Andersen M N, Jacobsen S E, Jensen C R. Physiological responses of potato (Solanum tuberosum L.) to partial root-zone drying: ABA signalling, leaf gas exchange, and water use efficiency. J Exp Bot, 2006, 57: 3727-3735. |
| [31] | Muñiz García M N, Cortelezzi J I, Fumagalli M, Capiati D A. Expression of the Arabidopsis ABF4 gene in potato increases tuber yield, improves tuber quality and enhances salt and drought tolerance. Plant Mol Biol, 2018, 98: 137-152. |
| [32] | Muñiz García M N, Giammaria V, Grandellis C, Téllez-Iñón M T, Ulloa R M, Capiati D A. Characterization of StABF1, a stress-responsive bZIP transcription factor from Solanum tuberosum L. that is phosphorylated by StCDPK2 in vitro. Planta, 2012, 235: 761-778. |
| [33] | Bai J P, Mao J, Yang H Y, Khan A, Fan A Q, Liu S Y, Zhang J L, Wang D, Gao H J, Zhang J L. Sucrose non-ferment 1 related protein kinase 2 (SnRK2) genes could mediate the stress responses in potato (Solanum tuberosum L.). BMC Genet, 2017, 18: 41. |
| [34] | 刘维刚, 唐勋, 付学, 张欢欢, 朱存兰, 张宁, 司怀军. 马铃薯抗旱性研究进展. 中国马铃薯, 2022, 36: 358-369. |
| Liu W G, Tang X, Fu X, Zhang H H, Zhu C L, Zhang N, Si H J. Research progress in drought tolerance of potato. Chin Potato J, 2022, 36: 358-369 (in Chinese with English abstract). | |
| [35] | Bouaziz D, Ayadi M, Bidani A, Rouis S, Nouri-Ellouz O, Jellouli R, Drira N, Gargouri-Bouzid R. A stable cytosolic expression of VH antibody fragment directed against PVY NIa protein in transgenic potato plant confers partial protection against the virus. Plant Sci, 2009, 176: 489-496. |
| [36] | Bouaziz D, Pirrello J, Charfeddine M, Hammami A, Jbir R, Dhieb A, Bouzayen M, Gargouri-Bouzid R. Overexpression of StDREB1 transcription factor increases tolerance to salt in transgenic potato plants. Mol Biotechnol, 2013, 54: 803-817. |
| [37] | Feller A, Machemer K, Braun E L, Grotewold E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J, 2011, 66: 94-116. |
| [38] | Li H M, Sun J Q, Xu Y X, Jiang H L, Wu X Y, Li C Y. The bHLH-type transcription factor AtAIB positively regulates ABA response in Arabidopsis. Plant Cell, 2007, 65: 655-665. |
| [39] | Yang T R, Yao S F, Hao L, Zhao Y Y, Lu W J, Xiao K. Wheat bHLH-type transcription factor gene TabHLH1 is crucial in mediating osmotic stresses tolerance through modulating largely the ABA-associated pathway. Plant Cell Rep, 2016, 35: 2309-2323. |
| [40] | Jiang Y Q, Yang B, Deyholos M K. Functional characterization of the Arabidopsis bHLH92 transcription factor in abiotic stress. Mol Genet Genomics, 2009, 282: 503-516. |
| [41] | Liu Y J, Ji X Y, Nie X G, Qu M, Zheng L, Tan Z L, Zhao H M, Huo L, Liu S N, Zhang B, Wang Y C. Arabidopsis AtbHLH112 regulates the expression of genes involved in abiotic stress tolerance by binding to their E-box and GCG-box motifs. New Phytol, 2015, 207: 692-709. |
| [42] | Khampheng B, 沈镭, 钟帅, 孙艳芝, 杨慧芹. 脯氨酸引发提高烟草种子和幼苗抗逆性及其与抗氧化系统的关系. 山西农业科学, 2019, 47(1): 39-48. |
| Khampheng B, Shen L, Zhong S, Sun Y Z, Yang H Q. Improving the antioxidant system and its stress resistance to tobacco seeds and seedling by proline priming. J Shanxi Agric Sci, 2019, 47(1): 39-48 (in Chinese with English abstract). | |
| [43] | Ji X Y, Nie X G, Liu Y J, Zheng L, Zhao H M, Zhang B, Huo L, Wang Y C. A bHLH gene from Tamarix hispida improves abiotic stress tolerance by enhancing osmotic potential and decreasing reactive oxygen species accumulation. Tree Physiol, 2016, 36: 193-207. |
| [1] | 郭飞翔, 李春霞, 周爽, 郭彬彬, 张均, 马超. 绿豆R2R3-MYB转录因子家族鉴定及其类黄酮合成调控基因的筛选[J]. 作物学报, 2025, 51(1): 117-133. |
| [2] | 祁稼民, 许春苗, 肖斌. 马铃薯TIFY基因家族的全基因组鉴定及表达分析[J]. 作物学报, 2024, 50(9): 2297-2309. |
| [3] | 杨煜琛, 靳雅荣, 骆金婵, 祝鑫, 李葳航, 贾纪原, 王小珊, 黄德均, 黄琳凯. 珍珠粟WD40基因家族鉴定及表达特征分析[J]. 作物学报, 2024, 50(9): 2219-2236. |
| [4] | 刘宸铭, 赵克勇, 悦曼芳, 赵延明, 吴忠义, 张春. 玉米转录因子ZmEREB180调控根系生长发育及耐逆的功能研究[J]. 作物学报, 2024, 50(8): 1920-1933. |
| [5] | 郭思语, 赵克勇, 代正罡, 邹华文, 吴忠义, 张春. 玉米N-乙酰转移酶ZmNAT1基因响应非生物胁迫的功能分析[J]. 作物学报, 2024, 50(8): 2001-2013. |
| [6] | 周洪源, 杨慧芹, 罗威, 石振明, 马玲. 马铃薯绿原酸调控因子的筛选与功能鉴定[J]. 作物学报, 2024, 50(7): 1740-1749. |
| [7] | 刘园园, 董建科, 应静文, 梅文祥, 程刚, 郭晶晶, 焦文标, 宋波涛. 利用野生种Solanum boliviense创制马铃薯抗寒种质[J]. 作物学报, 2024, 50(6): 1384-1393. |
| [8] | 刘震, 陈丽敏, 李志涛, 朱金勇, 王玮璐, 齐喆颖, 姚攀锋, 毕真真, 孙超, 白江平, 刘玉汇. 马铃薯ARM基因家族的全基因组鉴定及表达分析[J]. 作物学报, 2024, 50(6): 1451-1466. |
| [9] | 赵娜, 刘宇曦, 张朝澍, 石瑛. 不同马铃薯淀粉含量差异的转录组学解析[J]. 作物学报, 2024, 50(6): 1503-1513. |
| [10] | 折萌, 郑登俞, 柯照, 吴忠义, 邹华文, 张中保. 玉米ZmGRAS13基因的克隆及功能研究[J]. 作物学报, 2024, 50(6): 1420-1434. |
| [11] | 李万, 李成, 程敏, 吴芳. 磷转运蛋白StPHO1.2提高马铃薯耐热性[J]. 作物学报, 2024, 50(2): 394-402. |
| [12] | 殷祥贞, 赵健鑫, 郝翠翠, 潘丽娟, 陈娜, 许静, 姜骁, 赵旭红, 王恩琪, 曹欢, 禹山林, 迟晓元. 花生转录因子基因AhWRI1的克隆及表达分析[J]. 作物学报, 2024, 50(12): 3155-3164. |
| [13] | 肖胜华, 陆妍, 李安子, 覃耀斌, 廖铭静, 闭兆福, 卓柑锋, 朱永红, 朱龙付. 棉花AP2/ERF转录因子GhTINY2负调控植株抗盐性的功能分析[J]. 作物学报, 2024, 50(1): 126-137. |
| [14] | 上官小霞, 杨琴莉, 李换丽. 基于CRISPR/Cas9的棉花GhbHLH71基因编辑突变体的分析[J]. 作物学报, 2024, 50(1): 138-148. |
| [15] | 王丽平, 王晓钰, 傅竞也, 王强. 玉米转录因子ZmMYB12提高植物抗旱性和低磷耐受性的功能鉴定[J]. 作物学报, 2024, 50(1): 76-88. |
|