欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (7): 1683-1696.doi: 10.3724/SP.J.1006.2022.14126
陈璐( ), 周淑倩, 李永新, 陈刚, 陆国权, 杨虎清()
), 周淑倩, 李永新, 陈刚, 陆国权, 杨虎清()
CHEN Lu(), ZHOU Shu-Qian, LI Yong-Xin, CHEN Gang, LU Guo-Quan, YANG Hu-Qing()
摘要:
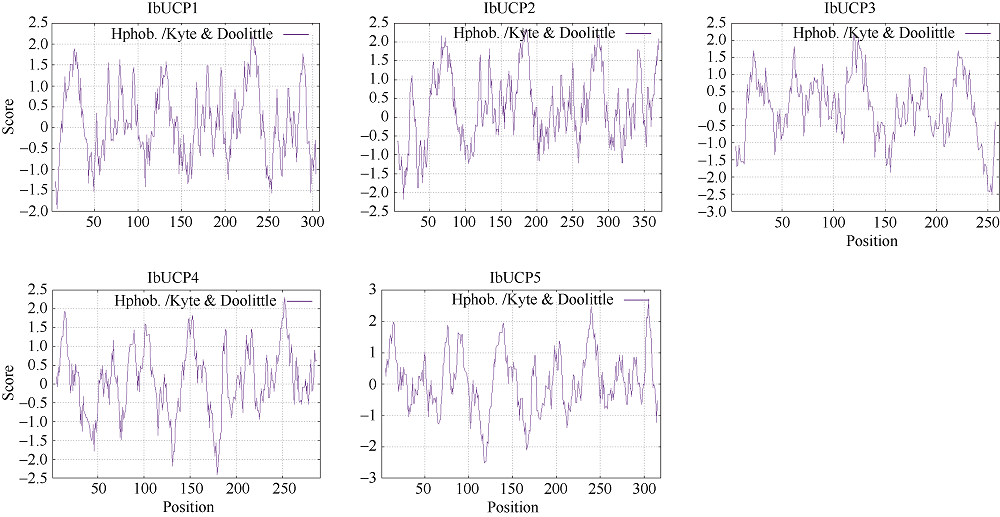
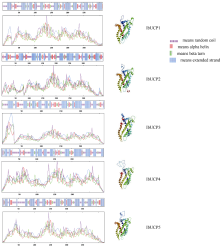
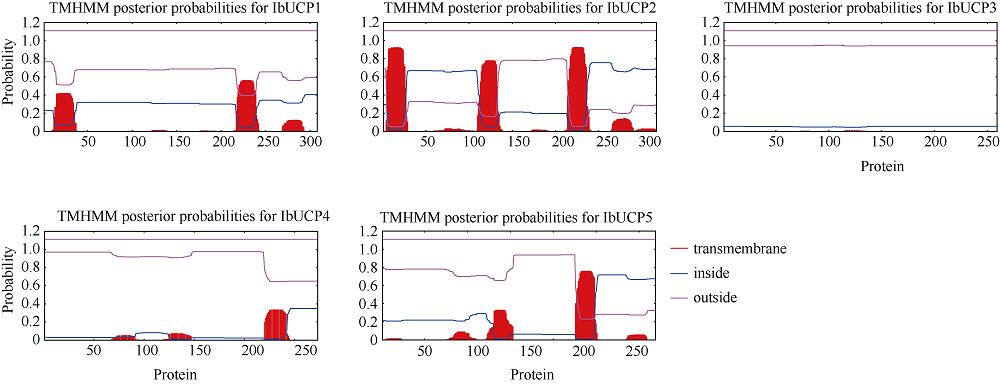
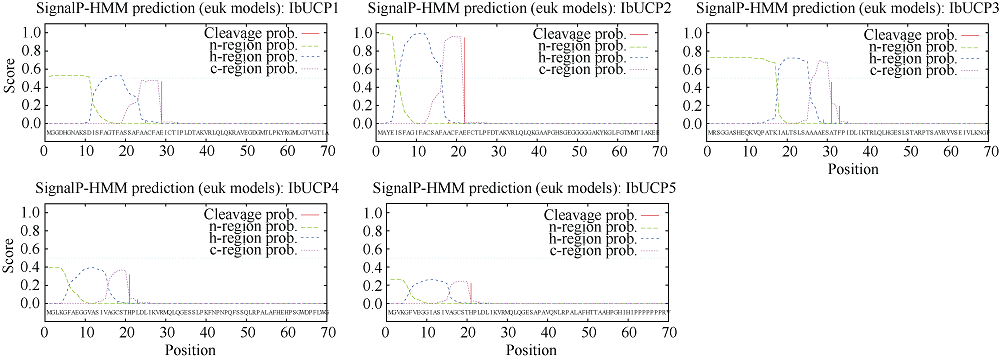
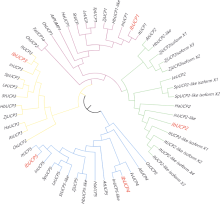
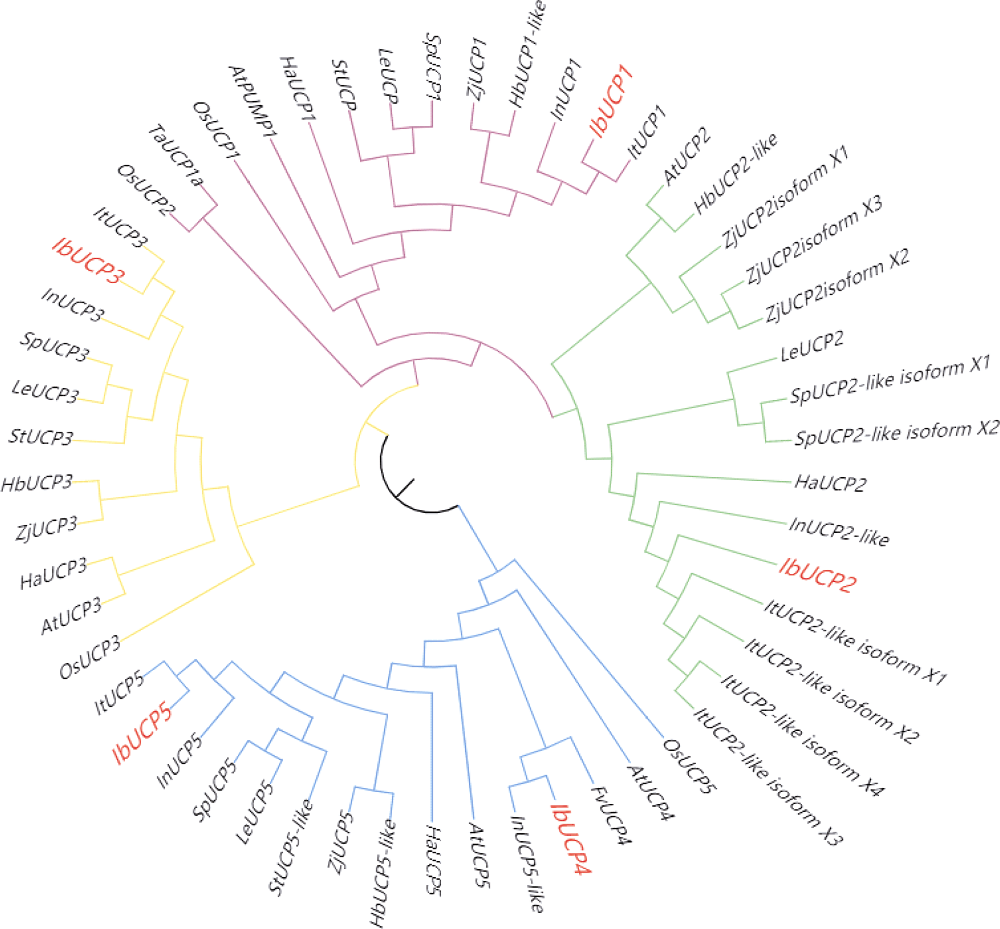
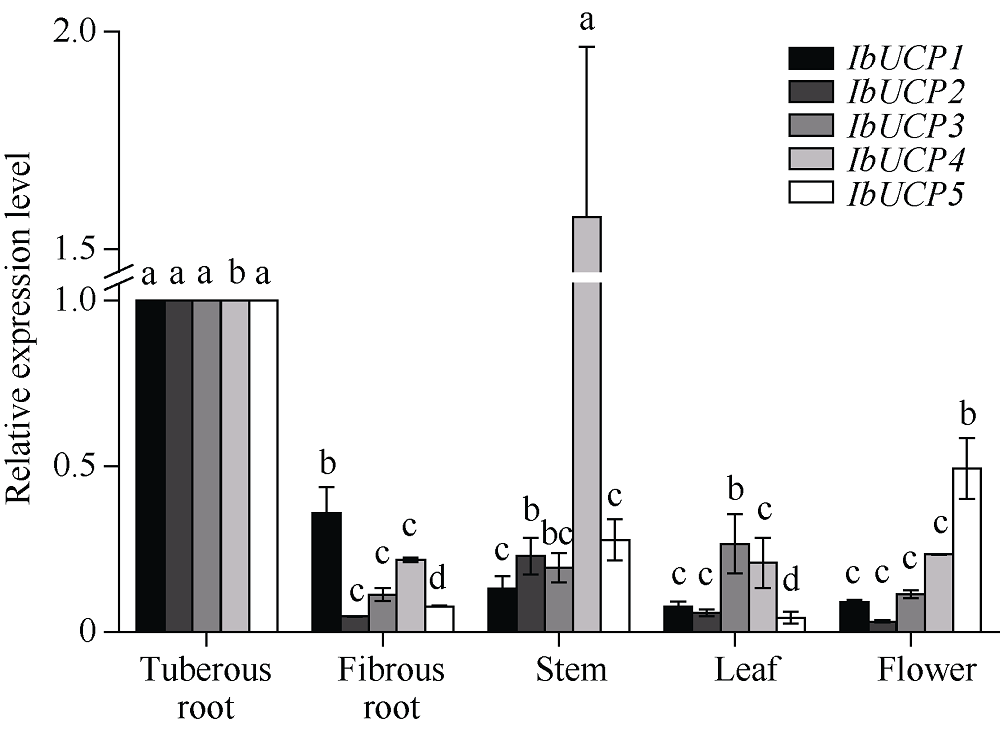
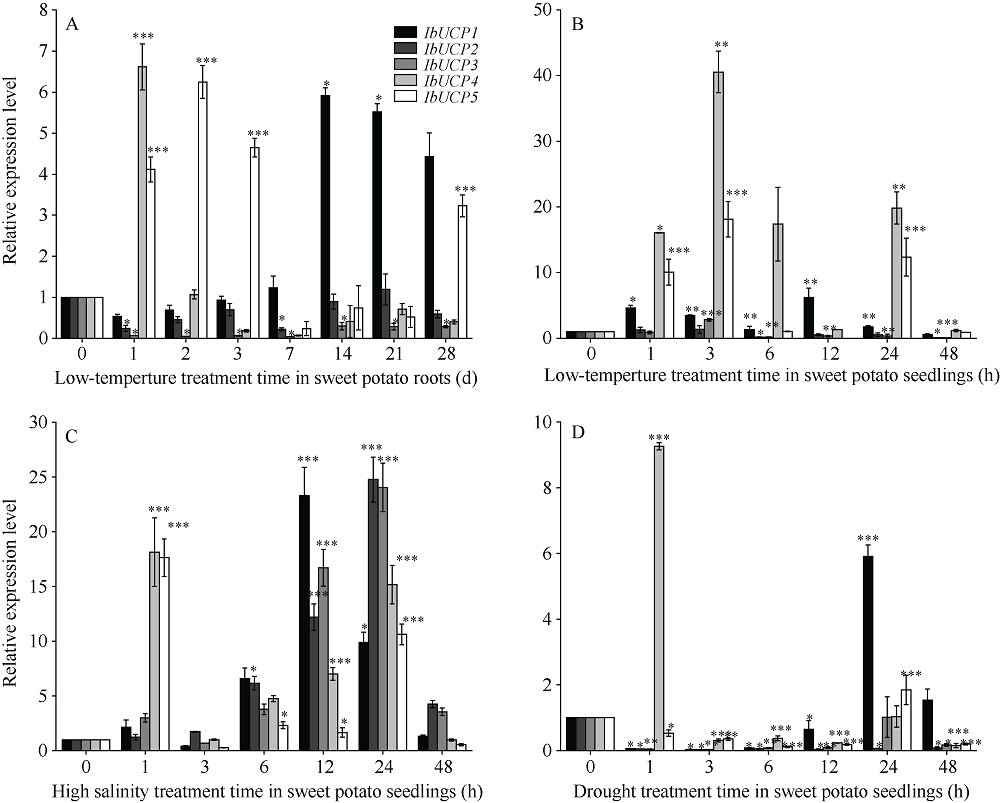
本研究旨在鉴定和分析甘薯(Ipomonea batatas (L.) Lam)解偶联蛋白(uncoupling protein, UCPs)基因家族成员, 探究其在甘薯不同组织中的表达特异性及其对低温(4℃)、高盐(NaCl)和干旱(PEG-6000)等胁迫的响应规律。结果发现, 甘薯UCP (IbUCP)含有5个家族基因, 分别将其命名为IbUCP1 (GenBank登录号为MW753000)、IbUCP2 (GenBank登录号为MW753004)、IbUCP3 (GenBank登录号为MW753001)、IbUCP4 (GenBank登录号为MW753002)和IbUCP5 (GenBank登录号为MW753003)。预测IbUCP的理论等电点为8.53~9.86, 含有261~375个的氨基酸残基; IbUCP定位于线粒体; IbUCP为亲水蛋白, 又属于线粒体载体蛋白超家族的成员, 其二级结构主要包括α-螺旋和无规则卷曲, 这与三级结构预测结果相符; IbUCP不存在跨膜螺旋结构和信号肽; IbUCP家族成员分为5个, 与三裂叶薯和牵牛花有较近的亲缘关系, 具有一定的保守性; 启动子预测发现, IbUCPs基因具有基本的转录元件以及一些信号响应元件、转录因子识别结合元件和逆境等响应顺式作用元件。表达分析显示, IbUCPs基因家族成员具有组织特异性, 其中IbUCP4在茎中表达最高, 其余IbUCPs均在块根中最高; IbUCPs基因家族成员中响应低温胁迫的有IbUCP1、IbUCP4和IbUCP5; IbUCPs基因家族对高盐胁迫均有响应; 在干旱的胁迫下, IbUCP1、IbUCP4和IbUCP5均有响应, 分别在不同的时间达到峰值。多种胁迫可调控IbUCPs的表达, 本研究为甘薯UCP基因的功能挖掘及甘薯抗逆品种筛选提供了一定的理论依据。
| [1] | Pradhan D M P, Mukherjee A, George J, Chakrabarti S K, Vimala B, Naskar S K, Sahoo B K, Samal S. High starch, beta carotene and anthocyanin rich sweet potato: ascent to future food and nutrition security in coastal and backward areas. Int J Trop Agric, 2015, 10: 9-22. |
| [2] | 徐飞, 袁澍, 梁厚果, 林宏辉. 交替氧化酶和解偶联蛋白在植物线粒体中的作用及其相互关系. 植物生理学通讯, 2009, 45(2): 105-110. |
| Xu F, Yuan S, Liang H G, Lin H H. The roles of alternative oxidase and uncoupling protein in plant mitochondria and their interrelationships. Plant Physiol Commun, 2009, 45(2): 105-110.(in Chinese with English abstract) | |
| [3] | 张海洋. 解偶联蛋白家族成员结构基础和功能机制的研究. 南京大学硕士学位论文, 江苏南京, 2015. |
| Zhang H Y. The Study on Structural Basis and Functinal Mechanismod Uncoupling Proteins. MS Thesis of Nanjing University, Nanjing, Jiangsu, China, 2015.(in Chinese with English abstract) | |
| [4] |
Sweetlove L J, Heazlewood J L, Herald V, Holtzapffel R H, Millar A H. The impact of oxidative stress on Arabidopsis mitochondria. J Plant, 2010, 32: 891-904.
doi: 10.1046/j.1365-313X.2002.01474.x |
| [5] |
Alscher R G, Donahue J L, Cramer C L. Reactive oxygen species and antioxidants: relationships in green cells. Physiol Plant, 1997, 100: 224-233.
doi: 10.1111/j.1399-3054.1997.tb04778.x |
| [6] |
Pastore D, Fratianni A, Di Pede S, Passarella S. Effects of fatty acids, nucleotides and reactive oxygen species on durum wheat mitochondria. FEBS Lett, 2000, 470: 88-92.
pmid: 10722851 |
| [7] |
Ricquier D, Kader J C. Mitochondrial protein alteration in active brown fat: a sodium dodecyl sulfate-polyacrylamide gel electrophoretic study. Biochem Biophys Res Commun, 1976, 73: 577-583.
doi: 10.1016/0006-291X(76)90849-4 |
| [8] |
Laloi M, Klein M, Riesmeier J W, Müller-Röber B, Fleury C, Bouillaud F, Ricquier D. A plant cold-induced uncoupling protein. Nature, 1997, 389: 135-136.
doi: 10.1038/38156 |
| [9] |
Maia I G, Benedetti C E, Leite A, Turcinelli S R, Arruda P. AtPUMP: an Arabidopsis gene encoding a plant uncoupling mitochondrial protein. FEBS Lett, 1998, 429: 403-406.
pmid: 9662458 |
| [10] |
Pastore D, Trono D, Laus M N, Di Fonzo N, Flagella Z. Possible plant mitochondria involvement in cell adaptation to drought stress. A case study: durum wheat mitochondria. J Exp Bot, 2007, 58: 195-210.
pmid: 17261694 |
| [11] |
Taylor N L, Heazlewood J L, Day D A, Millar A H. Differential impact of environmental stresses on the pea mitochondrial proteome. Mol Cell Proteomics, 2005, 4: 1122-1133.
doi: 10.1074/mcp.M400210-MCP200 |
| [12] | 刘自梅. 番茄线粒体解偶联蛋白基因(LeUCP)沉默对番茄光合作用, 呼吸作用及抗逆性的影响. 浙江大学硕士学位论文, 浙江杭州, 2011. |
| Liu Z M. Effects of Mitochondrial Uncoupled Protein Gene Silencing on Photosynthesis, Respiration and Stress Resistance of Tomato. MS Thesis of Zhejiang University, Hangzhou, Zhejiang, China, 2011.(in Chinese with English abstract) | |
| [13] |
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods an Arabidopsis mitochondrial uncoupling protein confers tolerance to drought and salt stress in transgenic tobacco plants. Mol Biol Evol, 2011, 28: 2731-2739.
doi: 10.1093/molbev/msr121 |
| [14] | 苏丽艳. 番茄SlETR6基因的克隆及非生物胁迫下的表达分析. 华北农学报, 2019, 34(1): 23-29. |
| Su L Y. Cloning and expression analysis of ethylene receptor gene SIETR6 in Solanum lycopersicum under abiotic stress. Acta Agric Boreali-Sin, 2019, 34(1): 23-29.(in Chinese with English abstract) | |
| [15] | 段奥其, 冯凯, 刘洁霞, 徐志胜, 熊爱生. 芹菜NAC转录因子基因AgNAC1的克隆及其对非生物胁迫的响应. 园艺学报, 2018, 45: 1125-1135. |
| Duan A Q, Feng K, Liu J X, Xu Z S, Xiong A S. Cloning and response to abiotic stress of NAC transcription gene AgNAC1 in Apium graveolens. Acta Hortic Sin, 2018, 45: 1125-1135 (in Chinese with English abstract) | |
| [16] |
Livak K, Schmittgen T. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25: 402-408.
doi: 10.1006/meth.2001.1262 pmid: 11846609 |
| [17] |
Barreto P, Couñago R M, Arruda P. Mitochondrial uncoupling protein-dependent signaling in plant bioenergetics and stress response. Mitochondrion, 2020, 53: 109-120.
doi: S1567-7249(19)30343-5 pmid: 32439620 |
| [18] |
Borecky J, Nogueira F, Oliveira K, Maia I G, Vercesi A E, Arruda P. The plant energy-dissipating mitochondrial systems: depicting the genomic structure and the expression profiles of the gene families of uncoupling protein and alternative oxidase in monocots and dicots. J Exp Bot, 2006, 57: 849-864.
pmid: 16473895 |
| [19] | Hourton-Cabassa C, Matos A R, Zachowski A, Moreau F. The plant uncoupling protein homologues: a new family of energy- dissipating proteins in plant mitochondria. J Plant Biochem Physiol, 2004, 42: 283-290. |
| [20] |
Ricquier D, Bouillaud F. The uncoupling protein homologues: UCP1, UCP2, UCP3, StUCP and AtUCP. Biochem J, 2000, 345: 161.
doi: 10.1042/bj3450161 |
| [21] |
Costa A, Nantes I L, Ježek P, Leite A, Vercesi A E. Plant uncoupling mitochondrial protein activity in mitochondria isolated from tomatoes at different stages of ripening. J Bioenerg Biomembr, 1999, 31: 527-533.
pmid: 10653480 |
| [22] |
Brandalise M, Maia I G, Boreck J, Vercesi A E, Arruda P. Overexpression of plant uncoupling mitochondrial protein in transgenic tobacco increases tolerance to oxidative stress. J Bioenerg Biomembr, 2003, 35: 203-209.
pmid: 13678271 |
| [23] |
Czobor Á, Hajdinák P, Németh B, Piros B, Németh Á, Szarka A. Comparison of the response of alternative oxidase and uncoupling proteins to bacterial elicitor induced oxidative burst. PLoS One, 2019, 14: e0210592.
doi: 10.1371/journal.pone.0210592 |
| [24] |
Calegario F F, Cosso R G, Fagian M M, Almeida F V, Jardim W F, Jezek P, Arruda P, Vercesi A E. Stimulation of potato tuber respiration by cold stress is associated with an increased capacity of both plant uncoupling mitochondrial protein (PUMP) and alternative oxidase. J Bioenerg Biomembr, 2003, 35: 211-220.
pmid: 13678272 |
| [25] |
Armstrong A F, Badger M R, Day D A, Barthet M M, Smith P M, Millar A H, Whelan J, Atkin O K. Dynamic changes in the mitochondrial electron transport chain underpinning cold acclimation of leaf respiration. Plant Cell Environ, 2010, 31: 1156-1169.
doi: 10.1111/j.1365-3040.2008.01830.x |
| [26] | Mizuno N, Sugie A, Kobayashi F, Takumi S. Mitochondrial alternative pathway is associated with development of freezing tolerance in common wheat. J Plant Biochem Physiol, 2008, 165: 462-467. |
| [27] | Ozawa K, Murayama S, Ai K U, Handa H. Overexpression of wheat mitochondrial uncoupling protein in rice plants confers tolerances to oxidative stresses promoted by exogenous hydrogen peroxide and low temperature. Mol Plant Breed, 2006, 18: 51-56. |
| [28] |
Popov V N, Eprintsev A T, Maltseva E V. Activation of genes encoding mitochondrial proteins involved in alternative and uncoupled respiration of tomato plants treated with low temperature and reactive oxygen species. Russ J Plant Physiol, 2011, 58: 914-920.
doi: 10.1134/S1021443711040091 |
| [29] |
Begcy K, Mariano E D, Mattiello L, Nunes A V, Mazzafera P, Maia I G, Menossi M. An Arabidopsis mitochondrial uncoupling protein confers tolerance to drought and salt stress in transgenic tobacco plants. PLoS One, 2011, 6: e23776.
doi: 10.1371/journal.pone.0023776 |
| [30] | 石晓雯. 甘薯逆境胁迫和花青素合成相关microRNA及其靶基因的鉴定和分析. 山西农业大学硕士学位论文, 山西太谷, 2018. |
| Shi X W. Identification and Analysis of microRNA and Their Target Genes Related to Anthocyanin Synthesis under Stress in Sweet Potato.MS Thesis of Shanxi Agricultural University, Taigu, Shanxi, China, 2018.(in Chinese with English abstract) | |
| [31] | 吴雨捷, 吴健, 王幼平, 孙勤富. WRKY转录因子在植物抗逆反应中的功能研究进展. 分子植物育种, 2020, 18: 7413-7422. |
| Wu Y J, Wu J, Wang Y P, Sun Q F. Advances in functional studies of WRKY transcription factors in plant adverse response. Mol Plant Breed, 2020, 18: 7413-7422.(in Chinese with English abstract) |
| [1] | 张天宇, 王越, 刘影, 周婷, 岳彩鹏, 黄进勇, 华营鹏. 油菜脯氨酸代谢基因家族的生物信息学分析与核心成员鉴定[J]. 作物学报, 2022, 48(8): 1977-1995. |
| [2] | 王沙沙, 黄超, 汪庆昌, 晁岳恩, 陈锋, 孙建国, 宋晓. 小麦籽粒大小相关基因TaGS2克隆及功能分析[J]. 作物学报, 2022, 48(8): 1926-1937. |
| [3] | 朱春权, 魏倩倩, 项兴佳, 胡文君, 徐青山, 曹小闯, 朱练峰, 孔亚丽, 刘佳, 金千瑜, 张均华. 褪黑素和茉莉酸甲酯基质育秧对水稻耐低温胁迫的调控作用[J]. 作物学报, 2022, 48(8): 2016-2027. |
| [4] | 白冬梅, 薛云云, 黄莉, 淮东欣, 田跃霞, 王鹏冬, 张鑫, 张蕙琪, 李娜, 姜慧芳, 廖伯寿. 不同花生品种芽期耐寒性鉴定及评价指标筛选[J]. 作物学报, 2022, 48(8): 2066-2079. |
| [5] | 解黎明, 姜仲禹, 柳洪鹃, 韩俊杰, 刘本奎, 王晓陆, 史春余. 甘薯发根分枝期适宜土壤水分促进块根糖供应和块根形成的研究[J]. 作物学报, 2022, 48(8): 2080-2087. |
| [6] | 郭家鑫, 鲁晓宇, 陶一凡, 郭慧娟, 闵伟. 棉花在盐碱胁迫下代谢产物及通路的分析[J]. 作物学报, 2022, 48(8): 2100-2114. |
| [7] | 颜佳倩, 顾逸彪, 薛张逸, 周天阳, 葛芊芊, 张耗, 刘立军, 王志琴, 顾骏飞, 杨建昌, 周振玲, 徐大勇. 耐盐性不同水稻品种对盐胁迫的响应差异及其机制[J]. 作物学报, 2022, 48(6): 1463-1475. |
| [8] | 陈松余, 丁一娟, 孙峻溟, 黄登文, 杨楠, 代雨涵, 万华方, 钱伟. 甘蓝型油菜BnCNGC基因家族鉴定及其在核盘菌侵染和PEG处理下的表达特性分析[J]. 作物学报, 2022, 48(6): 1357-1371. |
| [9] | 王霞, 尹晓雨, 于晓明, 刘晓丹. 干旱锻炼对B73自交后代当代干旱胁迫记忆基因表达及其启动子区DNA甲基化的影响[J]. 作物学报, 2022, 48(5): 1191-1198. |
| [10] | 雷新慧, 万晨茜, 陶金才, 冷佳俊, 吴怡欣, 王家乐, 王鹏科, 杨清华, 冯佰利, 高金锋. 褪黑素与2,4-表油菜素内酯浸种对盐胁迫下荞麦发芽与幼苗生长的促进效应[J]. 作物学报, 2022, 48(5): 1210-1221. |
| [11] | 晋敏姗, 曲瑞芳, 李红英, 韩彦卿, 马芳芳, 韩渊怀, 邢国芳. 谷子糖转运蛋白基因SiSTPs的鉴定及其参与谷子抗逆胁迫响应的研究[J]. 作物学报, 2022, 48(4): 825-839. |
| [12] | 巫燕飞, 胡琴, 周棋, 杜雪竹, 盛锋. 水稻延伸因子复合体家族基因鉴定及非生物胁迫诱导表达模式分析[J]. 作物学报, 2022, 48(3): 644-655. |
| [13] | 靳容, 蒋薇, 刘明, 赵鹏, 张强强, 李铁鑫, 王丹凤, 范文静, 张爱君, 唐忠厚. 甘薯Dof基因家族挖掘及表达分析[J]. 作物学报, 2022, 48(3): 608-623. |
| [14] | 丁红, 徐扬, 张冠初, 秦斐斐, 戴良香, 张智猛. 不同生育期干旱与氮肥施用对花生氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 695-703. |
| [15] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
|