欢迎访问作物学报,今天是
作物学报 ›› 2023, Vol. 49 ›› Issue (8): 2077-2087.doi: 10.3724/SP.J.1006.2023.23062
王兴荣1( ), 张彦军1, 涂奇奇2, 龚佃明2,*(), 邱法展2,*()
), 张彦军1, 涂奇奇2, 龚佃明2,*(), 邱法展2,*()
WANG Xing-Rong1(), ZHANG Yan-Jun1, TU Qi-Qi2, GONG Dian-Ming2,*(), QIU Fa-Zhan2,*()
摘要:
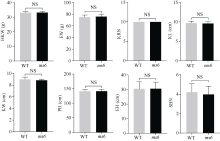
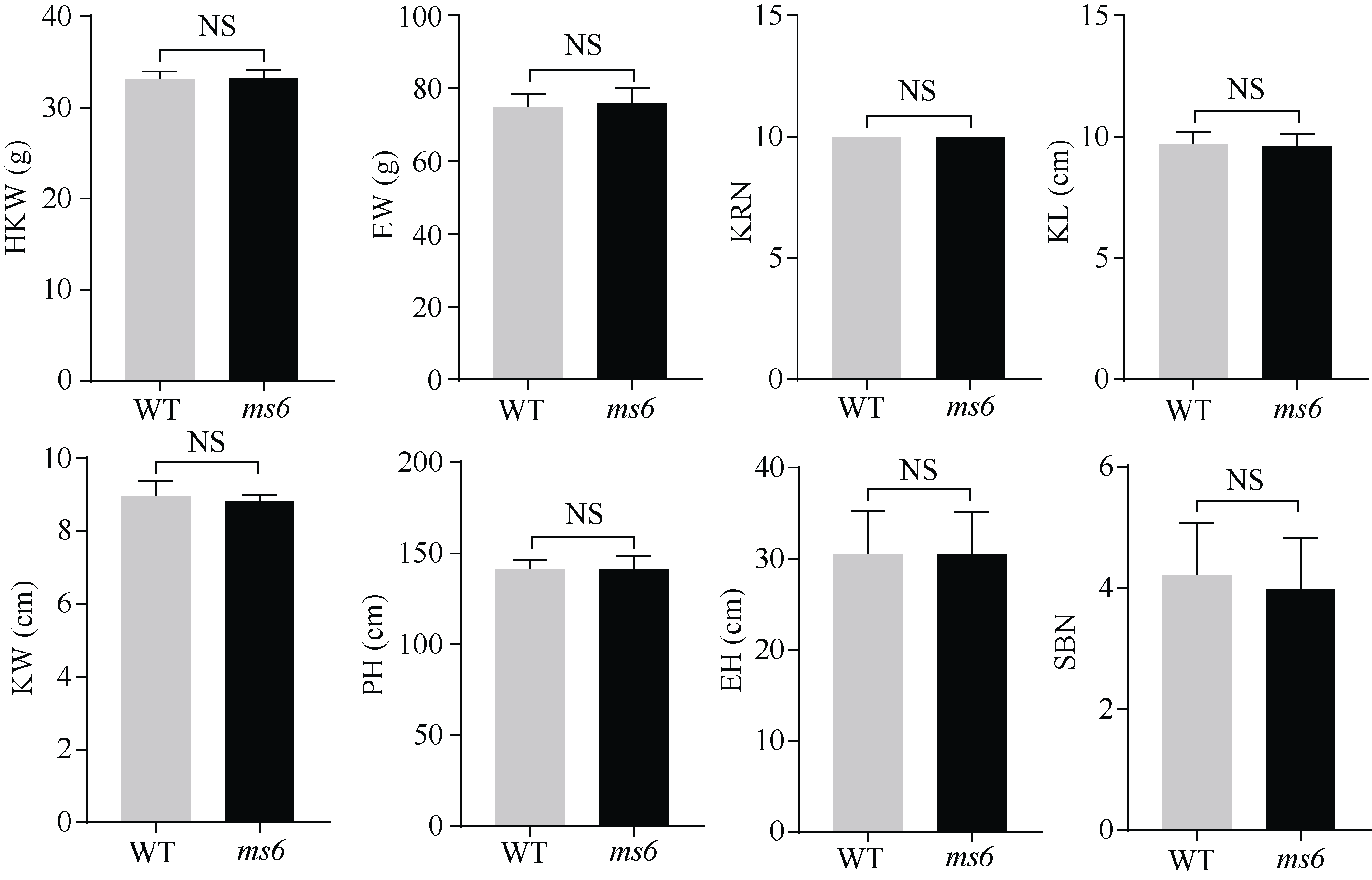
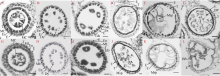
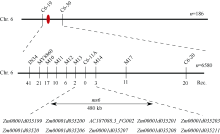
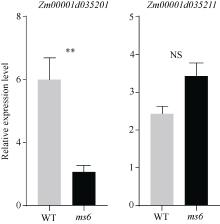
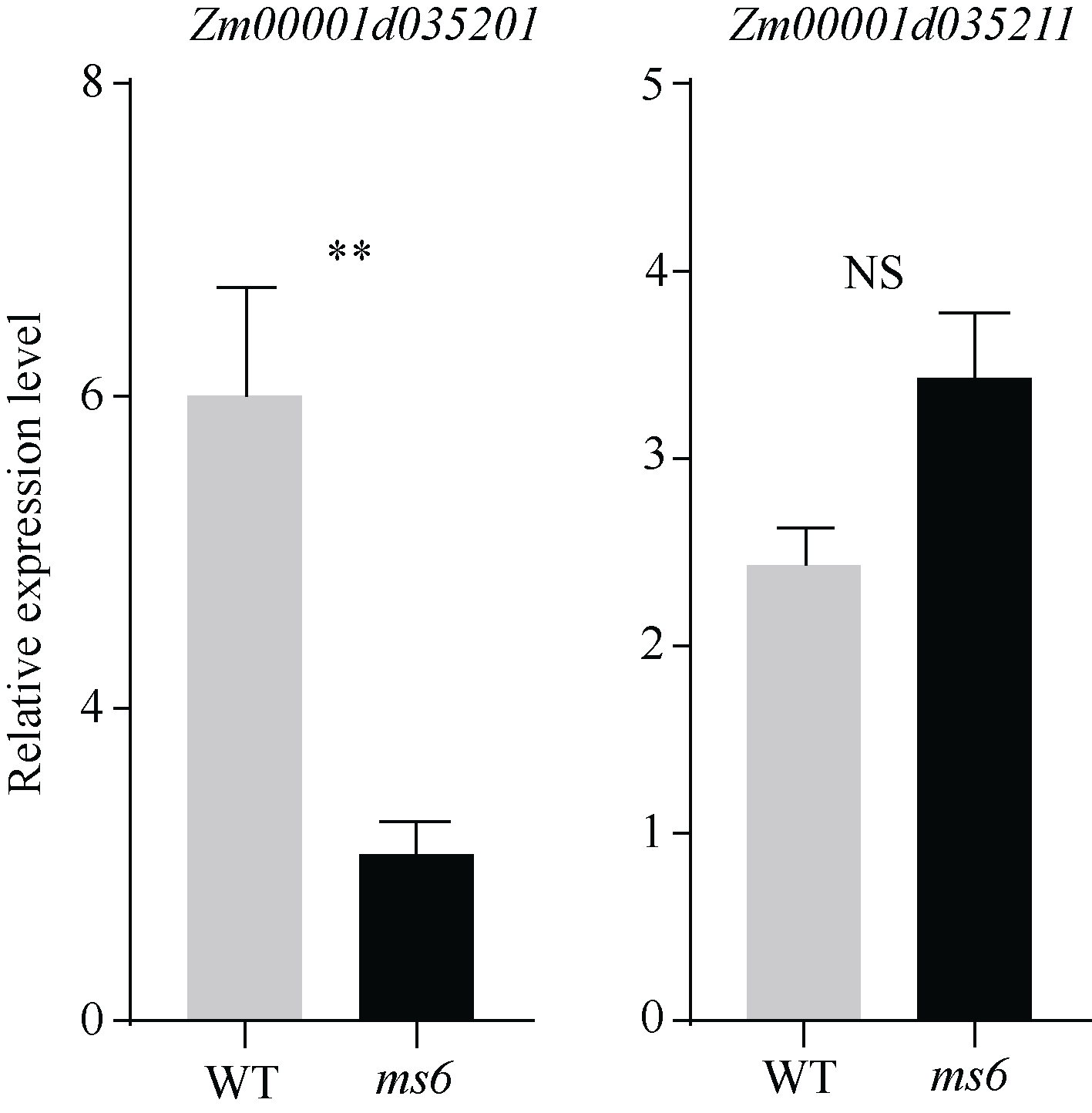
玉米雄性不育基因的定位、克隆和功能机理研究, 不仅能够加深我们对玉米雄花生长发育的分子调控机理的认识, 而且能够有效推动雄性不育技术体系的发展及在玉米育种和种子生产中的运用。本研究以玉米自交系Mo17为野生型背景材料, 经EMS诱变获得了一个玉米雄性不育突变体, 命名为ms6 (male sterile 6)。表型鉴定结果表明, ms6突变体植株能够正常抽雄, 但雄花颖壳不能正常开裂和散粉, 花粉粒干瘪, 表现为无花粉型不育。同时, ms6与Mo17野生型(wild type, WT)在株型、穗部性状以及籽粒粒形等相关性状上无显著差异, 说明该基因突变后, 仅影响植株的育性, 而不影响其他农艺性状。细胞学观察显示, ms6不育突变体的小孢子发育晚期出现异常, 表现为绒毡层细胞提前降解, 小孢子不能进行有丝分裂并逐渐裂解。扫描电镜观察表明, ms6花药外壁皱缩, 内壁无完整的花粉粒, 无乌氏小体的存在。遗传学分析表明, ms6突变性状受1对隐性核基因控制。以ms6 × B73 F2遗传定位群体, 利用全基因组约200对多态性SSR分子标记, 结合表型与基因型连锁分析, 将ms6初定位于玉米6号染色体C6-19与C6-30两个标记之间, 进一步利用区间内10对新开发的多态性标记, 最终将ms6定位在分子标记M13~M14之间约480 kb的区间范围内。转录组测序结合qRT-PCR试验验证结果, 初步将Zm00001d035201确定为ms6的关键候选基因。Zm00001d035201基因编码一个酸性核糖体蛋白。本研究结果为ms6后续基因功能的研究打下了坚实的基础。同时ms6作为一个新的核不育突变体也为将来玉米新型核不育基因的生产应用提供了重要材料支持。
| [1] | 吴锁伟, 张丹凤, 方才臣, 邓联武, 万向元. 玉米高效农杆菌转化体系的研究进展及其影响因素分析. 玉米科学, 2012, 20(5): 59-64. |
| Wu S W, Zhang D F, Fang C C, Deng L W, Wan X Y. Advances and major influencing factors of high-efficient agrobacterium- mediated genetic transformation in maize. J Maize Sci, 2012, 20(5): 59-64. (in Chinese with English abstract) | |
| [2] | 王保明, 陈永忠, 李红波, 莫华, 黄露波. 植物雄性不育的机制及应用研究进展. 河南农业科学, 2019, 48(5): 1-9. |
| Wang B M, Chen Y Z, Li H B, Mo H, Huang L B. Progress of mechanism of male sterility of plants and its application. J Henan Agric Sci, 2019, 48(5):1-9. (in Chinese with English abstract) | |
| [3] |
Wasiak M, Niedziela A, Woś H, Pojmaj M, Bednarek P T. Genetic mapping of male sterility and pollen fertility QTLs in triticale with sterilizing Triticum timopheevii cytoplasm. J Appl Genet, 2021, 62: 59-71.
doi: 10.1007/s13353-020-00595-z |
| [4] | 孙小媛, 王一帆, 王韫慧, 蔺佳雨, 李金红, 丘远涛, 方小龙, 孔凡江, 李美娜. 大豆细胞核雄性不育基因研究进展. 遗传, 2021, 43: 52-65. |
| Sun X Y, Wang Y F, Wang Y H, Lin J Y, Li J H, Qiu Y T, Fang X L, Kong F J, Li M N. Progress on genic male sterility gene in soybean. Hereditas, 2021, 43: 52-65. (in Chinese with English abstract) | |
| [5] |
Goldberg R B, Beals T P, Sanders P M. Anther development: basic principles and practical applications. Plant Cell, 1993, 5: 1217-1229.
doi: 10.1105/tpc.5.10.1217 pmid: 8281038 |
| [6] |
Chen X Y, Zhang H, Sun H Y, Luo H B, Zhao L, Dong Z B, Yan S S, Zhao C, Liu R Y, Xu C Y, Li S, Chen H B, Jin W W. IRREGULAR POLLEN EXINE1 is a novel factor in anther cuticle and pollen exine formation. Plant Physiol, 2017, 173: 307-325.
doi: 10.1104/pp.16.00629 |
| [7] |
Zhang D B, Wilson Z A. Stamen specification and anther development in rice. Chin Sci Bull, 2009, 54: 2342-2353.
doi: 10.1007/s11434-009-0348-3 |
| [8] | 田有辉, 万向元. 玉米花药发育的细胞生物学与分子遗传学的研究方法. 中国生物工程杂志, 2018, 38(1): 88-89. |
| Tian Y H, Wang X Y. The methods of cell biology and molecular genetics of maize anther development. Chin J Biotechnol, 2018, 38(1): 88-89. (in Chinese with English abstract) | |
| [9] |
Wan X Y, Wu S W, Li Z W, Dong Z Y, An X L, Ma B, Tian Y H, Li J P. Maize genic male-sterility genes and their applications in hybrid breeding: progress and perspectives. Mol Plant, 2019, 12: 321-342.
doi: S1674-2052(19)30020-6 pmid: 30690174 |
| [10] |
Zhang D F, Wu S W, An X L, Xie K, Dong Z Y, Zhou Y, Xu L W, Fang W, Liu S S, Liu S S, Zhu T T, Li J P, Rao L Q, Zhao J R, Wan X Y. Construction of a multicontrol sterility system for a maize male-sterile line and hybrid seed production based on the ZmMs7 gene encoding a PHD-finger transcription factor. Plant Biotechnol J, 2018, 16: 459-471.
doi: 10.1111/pbi.2018.16.issue-2 |
| [11] | Nan G L, Zhai J, Arikit S, Morrow D, Fernandes J, Mai L, Nguyen N, Meyers B C, Walbot V. MS23, a master basic helix-loop-helix factor, regulates the specification and development of the tapetum in maize. Development, 2017, 144: 163-172. |
| [12] |
Moon J, Skibbe D, Timofejeva L, Wang C J, Kelliher T, Kremling K, Walbot V, Cande W Z. Regulation of cell divisions and differentiation by MALE STERILITY32 is required for anther development in maize. Plant J, 2013, 76: 592-602.
doi: 10.1111/tpj.2013.76.issue-4 |
| [13] |
Vernoud V, Laigle G, Rozier F, Meeley R B, Perez P, Rogowsky P M. The HD-ZIP IV transcription factor OCL4 is necessary for trichome patterning and anther development in maize. Plant J, 2009, 59: 883-894.
doi: 10.1111/tpj.2009.59.issue-6 |
| [14] |
Wang D, Adams C M, Fernandes J F, Egger R L, Walbot V.A low molecular weight proteome comparison of fertile and male sterile 8 anthers of Zea mays. Plant Biotechnol J, 2012, 10: 925-935.
doi: 10.1111/j.1467-7652.2012.00721.x pmid: 22748129 |
| [15] |
Wang Y B, Liu D C, Tian Y H, Wu S W, An X L, Dong Z Y, Zhang S M, Bao J X, Li Z W, Li J P, Wan X Y. Map-based cloning, phylogenetic, and microsynteny analyses of ZmMs20 gene regulating male fertility in maize. Int J Mol Sci, 2019, 20: 1411.
doi: 10.3390/ijms20061411 |
| [16] |
Djukanovic V, Smith J, Lowe K, Yang M Z, Gao H R, Jones S, Nicholson M G, West A, Lape J, Bidney D, Carl Falco S, Jantz D, Alexander Lyznik L. Male-sterile maize plants produced by targeted mutagenesis of the cytochrome P450-like gene (MS26) using a re-designed I-CreI homing endonuclease. Plant J, 2013, 76: 888-899.
doi: 10.1111/tpj.12335 |
| [17] |
An X L, Dong Z Y, Tian Y H, Xie K, Wu S W, Zhu T T, Zhang D F, Zhou Y, Niu C F, Ma B, Hou Q C, Bao J X, Zhang S M, Li Z W, Wang Y B, Yan T W, Sun X J, Zhang Y W, Li J P, Wan X Y. ZmMs30 encoding a novel GDSL lipase is essential for male fertility and valuable for hybrid breeding in maize. Mol Plant, 2019, 12: 343-359.
doi: 10.1016/j.molp.2019.01.011 |
| [18] |
Xie K, Wu S W, Li Z W, Zhou Y, Zhang D F, Dong Z Y, An X L, Zhu T T, Zhang S M, Liu S S, Li J P, Wan X Y. Map-based cloning and characterization of Zea mays male sterility 33 (ZmMs33) gene, encoding a glycerol-3-phosphate acyltransferase. Theor Appl Genet, 2018, 131: 1363-1378.
doi: 10.1007/s00122-018-3083-9 pmid: 29546443 |
| [19] |
Fox T, DeBruin J, Haug Collet K, Trimnell M, Clapp J, Leonard A, Li B, Scolaro E, Collinson S, Glassman K, Miller M, Schussler J, Dolan D, Liu L, Gho C, Albertsen M, Loussaert D, Shen B. A single point mutation in Ms44 results in dominant male sterility and improves nitrogen use efficiency in maize. Plant Biotechnol J, 2017, 15: 942-952.
doi: 10.1111/pbi.2017.15.issue-8 |
| [20] |
Cigan A M, Unger E, Xu R J. Phenotypic complementation of ms45 maize requires tapetal expression of MS45. Sex Plant Reprod, 2001, 14: 135-142.
doi: 10.1007/s004970100099 |
| [21] |
Tian Y H, Xiao S L, Liu J, Somaratne Y, Zhang H, Wang M M, Zhang H R, Zhao L, Chen H B. MALE STERILE6021 (MS6021) is required for the development of anther cuticle and pollen exine in maize. Sci Rep, 2017, 7: 16736.
doi: 10.1038/s41598-017-16930-0 |
| [22] |
Somaratne Y, Tian Y H, Zhang H, Wang M M, Huo Y Q, Cao F G, Zhao L, Chen H B. ABNORMAL POLLEN VACUOLATION1 (APV1) is required for male fertility by contributing to anther cuticle and pollen exine formation in maize. Plant J, 2017, 90: 96-110.
doi: 10.1111/tpj.2017.90.issue-1 |
| [23] |
Wang C J, Nan G L, Kelliher T, Timofejeva L, Vernoud V, Golubovskaya I N, Harper L, Egger R, Walbot V, Cande W Z. Maize multiple archesporial cells 1 (mac1), an ortholog of rice TDL1A, modulates cell proliferation and identity in early anther development. Development, 2012, 139: 2594-2603.
doi: 10.1242/dev.077891 |
| [24] | 曹双河, 张相岐, 张爱民. 光(温)敏雄性不育的调控机理和分子遗传学研究进展. 植物学通报, 2005, 22: 19-26. |
| Cao S H, Zhang X Q, Zhang A M. Review of the molecular regulation mechanism and genetics of photoperiod- and/or thermosensitive male sterility. Chin Bull Bot, 2005, 22: 19-26. (in Chinese with English abstract) | |
| [25] | 谢潮添, 杨延红, 葛丽丽, 王瑞, 田惠桥. 白菜核雄性不育花药超微结构的研究. 实验生物学报, 2005, 38: 501-512. |
| Xie C T, Yang Y H, Ge L L, Wang R, Tian H Q. The study on ultrastructure of anther of male sterility in Chinese cabbage. Acta Biol Exp Sin, 2005, 38: 501-512. (in Chinese with English abstract) | |
| [26] |
Vidakovic M B, Vancetovic J, Vidakovic M. A new search for restorer cytoplasm: the restorer cytoplasm for the gene ms10 most probably does not exist in maize. J Hered, 2002, 93: 444-447.
pmid: 12642646 |
| [27] | 王超, 安学丽, 张增为, 杨青, 饶力群, 陈信波, 方才臣, 万向元. 植物隐性核雄性不育基因育种技术体系的研究进展与展望. 植物生物工程杂志, 2013, 33(10): 124-130. |
| Wang C, An X L, Zhang Z W, Yang Q, Rao L Q, Chen X B, Fang C C, Wan X Y. Research progress and prospect of plant recessive nuclear male sterile gene breeding technology system. Chin J Biotechnol, 2013, 33(10): 124-130. (in Chinese with English abstract) | |
| [28] |
An X L, Ma B, Duan M J, Dong Z Y, Liu R G, Yuan D Y, Hou Q C, Wu S W, Zhang D F, Liu D C, Yu D, Zhang Y W, Xie K, Zhu T T, Li Z W, Zhang S M, Tian Y H, Liu C, Li J P, Yuan L P, Wan X Y. Molecular regulation of ZmMs7 required for maize male fertility and development of a dominant male-sterility system in multiple species. Proc Natl Acad Sci USA, 2020, 117: 23499-23509.
doi: 10.1073/pnas.2010255117 |
| [1] | 艾蓉, 张春, 悦曼芳, 邹华文, 吴忠义. 玉米转录因子ZmEREB211对非生物逆境胁迫的应答[J]. 作物学报, 2023, 49(9): 2433-2445. |
| [2] | 黄钰杰, 张啸天, 陈会丽, 王宏伟, 丁双成. 玉米ZmC2s基因家族鉴定及ZmC2-15耐热功能分析[J]. 作物学报, 2023, 49(9): 2331-2343. |
| [3] | 杨文宇, 吴成秀, 肖英杰, 严建兵. 基于Adaptive Lasso的两阶段全基因组关联分析方法[J]. 作物学报, 2023, 49(9): 2321-2330. |
| [4] | 唐杰, 龙湍, 吴春瑜, 李新鹏, 曾翔, 吴永忠, 黄培劲. 水稻OsGMS2基因的鉴定及其核不育系种子繁殖体系构建[J]. 作物学报, 2023, 49(8): 2025-2038. |
| [5] | 白岩, 高婷婷, 卢实, 郑淑波, 路明. 近四十年来我国玉米大品种的历史沿革与发展趋势[J]. 作物学报, 2023, 49(8): 2064-2076. |
| [6] | 王娟, 徐相波, 张茂林, 刘铁山, 徐倩, 董瑞, 刘春晓, 关海英, 刘强, 汪黎明, 何春梅. 一个新的玉米Miniature1基因等位突变体的鉴定与遗传分析[J]. 作物学报, 2023, 49(8): 2088-2096. |
| [7] | 韦金贵, 郭瑶, 柴强, 殷文, 樊志龙, 胡发龙. 水氮减量密植玉米的产量及产量构成[J]. 作物学报, 2023, 49(7): 1919-1929. |
| [8] | 李荣, 勉有明, 侯贤清, 李培富, 王西娜. 施氮对还田秸秆腐解及养分释放、土壤肥力与玉米产量的影响[J]. 作物学报, 2023, 49(7): 2012-2022. |
| [9] | 梅秀鹏, 赵子堃, 贾欣瑶, 白洋, 李梅, 甘宇玲, 杨秋悦, 蔡一林. 热诱导转录因子ZmNF-YC13调控热胁迫应答基因提高玉米耐热性[J]. 作物学报, 2023, 49(7): 1747-1757. |
| [10] | 常丽娟, 梁晋刚, 宋君, 刘文娟, 付成平, 代晓航, 王东, 魏超, 熊梅. 转基因玉米ND207转化事件特异性定性PCR检测方法及其标准化[J]. 作物学报, 2023, 49(7): 1818-1828. |
| [11] | 林孝欣, 黄明江, 韦祎, 朱洪慧, 王子怡, 李忠成, 庄慧, 李彦羲, 李云峰, 陈锐. 水稻籽粒伸长突变体lgdp的鉴定与基因定位[J]. 作物学报, 2023, 49(6): 1699-1707. |
| [12] | 张振博, 贾春兰, 任佰朝, 刘鹏, 赵斌, 张吉旺. 氮磷配施对夏玉米产量和叶片衰老特性的影响[J]. 作物学报, 2023, 49(6): 1616-1629. |
| [13] | 刘佳, 邹晓悦, 马继芳, 王永芳, 董志平, 李志勇, 白辉. 谷子MAPK家族成员的鉴定及其对生物胁迫的响应分析[J]. 作物学报, 2023, 49(6): 1480-1495. |
| [14] | 李璐璐, 明博, 高尚, 谢瑞芝, 王克如, 侯鹏, 薛军, 李少昆. 不同熟期玉米品种籽粒田间脱水特征差异性分析[J]. 作物学报, 2023, 49(6): 1643-1652. |
| [15] | 王玉珑, 于爱忠, 吕汉强, 吕奕彤, 苏向向, 王鹏飞, 柴健. 绿洲灌区麦后复种绿肥并还田对翌年玉米根系性状及水分利用效率的影响[J]. 作物学报, 2023, 49(5): 1350-1362. |
|